

Subfamily: Parvovirinae
Genus: Dependoparvovirus
Distinguishing features
Viruses in genus Dependoparvovirus are distinguished by their position in the phylogenetic tree (Figure 6B. Parvoviridae), where they share a deep branch with members of genera Copiparvovirus, Erythroparvovirus and Tetraparvovirus, and by standard parvovirus sequence identity criteria. Although all genomes are homotelomeric and show high sequence identity, members of one species, Anseriform dependoparvovirus 1, have many atypical characteristics.
The type species, Adeno-associated dependoparvovirus A, includes adeno-associated viruses (AAVs) 1–4 and 6–13, which infect primates and have genomes flanked by terminal repeat sequences (TRs) of ~145 nt, ending in T-shaped ~125 nt hairpin telomeres. These viruses are defective, only undergoing productive replication in cells that are co-infected with another DNA virus, most commonly an adenovirus, which modifies the host environment in multiple ways that are essential to enable the AAV life cycle: by themselves AAVs are not associated with any known pathology. Under certain conditions, for example in the presence of DNA damaging agents, limited AAV replication can also be detected in the absence of helper viruses. Six other species (Adeno-associated dependoparvovirus B, Avian dependoparvovirus 1, Chiropteran dependoparvovirus 1, Pinniped dependoparvovirus 1, Squamate dependoparvovirus 1 and Squamate dependoparvovirus 2) in this genus contain viruses with genomes that resemble those of AAV and are either known to be helper-virus dependent or are thought likely to be. Viruses from these species infect a wide range of mammalian, avian or reptilian hosts. In contrast, viruses in species Anseriform dependoparvovirus 1 are helper-virus independent, have genomes flanked by extremely long TRs (~440–460 nt) of which the distal ~407–420 nt fold into hairpins, have atypical gene expression strategies, and are highly pathogenic in naive populations of ducks and geese, especially in young birds, causing epidemics of a variable syndrome collectively dubbed Derzsy’s disease (Zádori et al., 1995).
Virion
See discussion under family description and Figure 1. Parvoviridae therein. AAV virion structures have been studied in great detail, in part because many of these viruses are being developed as vectors for the clinical treatment of genetic disorders involving a broad range of differentiated human cell types. X-ray and cryo-EM image reconstructions have been used to determine the structures of AAV variants 1-9 (sometimes called “serotypes” and including viruses from two different species, Adeno-associated dependoparvovirus A and Adeno-associated dependoparvovirus B), recombinant AAV vectors carrying foreign genetic information (rAAVs), capsid mutants, or any of the above complexed with specific antibodies, glycans or other receptors, at high resolution. While it would be impractical to review this field in the current forum, the following few papers illustrate some areas of interest (Agbandje-McKenna and Kleinschmidt 2011, Tseng et al., 2015, Huang et al., 2016, Xie et al., 2017, Crispino et al., 2017, Kanaan et al., 2017, Tse et al., 2017, Bennett et al., 2018).
Genome organization and replication
Dependoparvovirus gene expression strategies follow at least three distinct patterns. The first pattern is best characterized in AAV2, the exemplar virus of type species Adeno-associated dependoparvovirus A, which has three transcriptional promoters, at 5, 19 and 40 map units that are expressed in temporal order, starting with P5 (Mouw and Pintel 2000). Since all transcripts are polyadenylated at a single site, 3 size classes of pre-mRNAs are generated (Figure 1. .Dependoparvovirus). However, like all members of genus Dependoparvovirus, each pre-mRNA class contains an intron in the middle of the genome which, when excised, utilizes a single splice donor site and either one of two splice acceptor sites. As a result, each pre-mRNA gives rise to three mRNAs, which are translated to create up to 3 distinct protein products depending on the efficiency and frame-shifting effects of the splice. In fact, in AAV2, the P5 transcripts give rise to two forms of the initiator protein, called Rep78 and Rep68, which respectively do and do not include a C-terminal peptide encoded downstream of the splice donor. Among all parvoviruses studied to date only the AAVs use a promoter at P19, which generates transcripts encoding truncated ancillary forms of the rep gene that lack the endonuclease but contain the helicase domain. Like their P5 counterparts, P19 transcripts give rise to two proteins depending on whether they are translated from spliced (Rep40) or unspliced (Rep52) mRNAs. P40 transcripts express the capsid sequences; these arise just upstream of the central intron, so that an ATG translation start site positioned between the two alternative splice acceptors allows them to differentially express either VP1 or VP2/3. Specifically, mRNAs encoding VP1 use the first acceptor site and translation initiates at the internal ATG, allowing expression of the VP1 specific region (VP1SR) and all downstream sequences, whereas mRNAs using the second acceptor initiate translation downstream of VP1SR, using alternative initiation sites (ACG and ATG) to give rise to VP2 and VP3, respectively. Unspliced P40 transcripts are relatively infrequent, and do not appear to be translated (Qiu and Pintel 2002).
|
|
|
Figure 1. .Dependoparvovirus. Genetic strategy of adeno-associated virus 2 (AAV2), genus Dependoparvovirus. The homotelomeric genome of AAV2 is shown as a single line flanked by boxed terminal repeats of 145 nt that terminate in 125 nt T-shaped hairpins (magnified relative to the rest of the genome). Regions encoding the replication initiator proteins, Rep68 and 78, are shaded red, those expressed in the capsid proteins are green, and those in the unique dependoparvovirus ancillary proteins Rep 40/52 and AAP (assembly-activating protein) are yellow. AAAAA indicates the single polyadenylation site. |
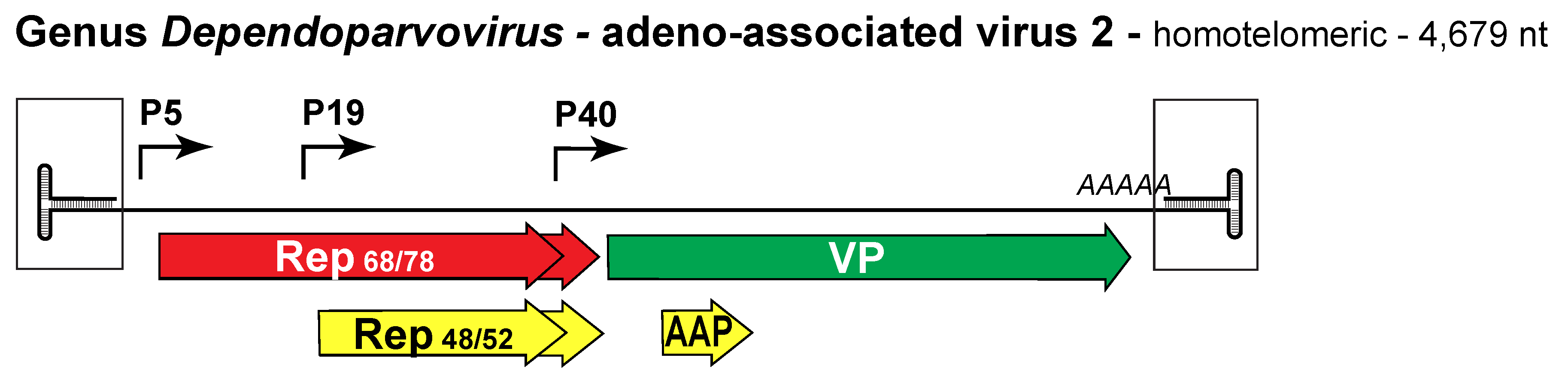
Loops within the Rep68 and 78 endonuclease domain mediate site-specific duplex DNA binding, recognizing a 22 bp sequence called the Rep binding element (RBE) that is present within both the terminal repeats (TRs) and the P5 promoter. During productive infection (in the presence of adenovirus) binding of these proteins to the RBE in the TR activates P5 while binding to the P5 RBE represses P5 transcription. However, both the TR and the P5 RBE transactivate the P19 and P40 promoters (Pereira et al., 1997).
A second transcription pattern, used by adeno-associated virus 5 (AAV5) and by several other non-primate AAV-like dependoparvoviruses also relies on 3 promoters (P7, P19 and P41) but is distinguished by the presence of an additional proximal polyadenylation site, (pA)p, derived from sequences positioned in the central intron, which is used to process all P7 and P19 transcripts. For AAV5 this means that only one form of the replication initiator protein, Rep 78, is expressed and while P19 still gives rise to both Rep52 and Rep40, these do not come from differentially spliced RNAs but are the products of differential translation initiation. (Qiu et al., 2006c, Qiu et al., 2006b, Ye and Pintel 2008).
The third characterized transcription pattern, used by members of species Anseriform dependoparvovirus 1 such as goose parvovirus (GPV, U25749), is a mixture of features seen in other parvoviruses but with some unique elements (Qiu et al., 2005). Like AAV5, these viruses use two polyadenylation sites and all the processed rep gene transcripts terminate at a position corresponding to the center of the genome at (pA)p, but in GPV some of the P9 transcripts are spliced prior to polyadenylation, causing two C-terminal variants of the initiator protein to be generated, called Rep1 and Rep1'. In other ways, the GPV expression strategy more closely resembles a pattern seen among members of genus Protoparvovirus: for example, GPV uses just two promoters, P9 and P42, and has an additional intron within Rep coding sequences that allows it to generate some spliced Rep2 mRNAs. However, these transcripts splice into the Rep1 ORF, rather than from it as seen in protoparvoviruses, and thus express the Rep1 helicase domain. The expression pattern of GPV is further complicated by a remarkable aspect of GPV RNA processing, in which nuclear processing of P9 pre-mRNAs controls the choice of translation start site subsequently used to initiate synthesis of Rep1 and Rep2, even though the intron sequences that determine this choice have already been excised from the cytoplasmic Rep2 mRNA (Li et al., 2009, Li and Pintel 2012). In silico analyses suggest that the reptilian dependoparvoviruses might use a similar splicing and polyadenylation pattern, although the presence of a promoter homologous to the P19 of AAV5 has been also predicted (Farkas et al., 2004, Pénzes et al., 2015).
The role of the GPV Rep2 protein remains uncertain, but the helicase activity of AAV ancillary proteins Rep40 and Rep52 has been shown to drive the packaging of progeny single-stranded DNA into pre-assembled viral capsids. These proteins oligomerize to form a capsid-tethered molecular motor that uses ATP hydrolysis to mediate strand translocation in a 3′-to-5′ direction into the capsid via one of its fivefold pores (King et al., 2001, Yoon-Robarts et al., 2004). Dependoparvoviruses also encode a ~23kDa ancillary assembly-activating protein (AAP) that is translated from a non-conventional start site (CTG) in an alternative ORF of the VP gene (Figure 1. .Dependoparvovirus). In AAV2 this protein acts as a chaperone, targeting newly synthesized capsid proteins to the nucleolus, where it promotes capsid assembly via interactions with VP C terminal peptides (Sonntag et al., 2010). AAVs 1 through 12 all encode AAPs, most of which can promote the assembly of capsids expressed by other AAV variants, but these proteins show significant variations. Thus, for example, there are variant-specific patterns of AAP-nucleolar association, some AAPs do not colocalize with assembled capsids, some assembled capsids are excluded from the nucleolus, and AAV4, AAV5 and AAV11 capsids are not dependent on AAP for assembly (Earley et al., 2017). This disparity was further explored by combining secondary structure analysis of the variants with mutagenesis and domain substitution, leading to the conclusion that AAPs have modular structures in which each module plays a distinct role and can potentially be engineered to carry out new functions (Tse et al., 2018). To date, capsid-assembly proteins with functions similar to AAP have not been identified in members of other genera although a small alternatively translated (SAT) protein with very different function does occupy a similar position and is expressed by an analogous mechanism in members of genus Protoparvovirus (Mészáros et al., 2017b).
Biology
See discussion under family description. Viruses in this genus were first reported in 1965 as ~20 nm particles contaminating adenovirus preparations (Atchison et al., 1965) and have been studied extensively during the intervening years making them among the best-characterized viruses in the family. AAVs appear to be fairly ubiquitous in humans and most other vertebrates but they have evolved a latent, defective life style, which means that there is no significant correlation between widespread AAV infection and any known disease (Kerr and Linden 2006, Zinn and Vandenberghe 2014). By the age of 10 >60% of children in the US have neutralizing antibodies to AAV serotypes 1–3, which often persist into adulthood.
In the late 1960s a contagious waterfowl disease swept across Europe, causing 70–100% mortality in goslings of <4 weeks old (Zádori et al., 2006). Originally given many different names, this infection was renamed Derzsy's disease in 1974 and both the goose virus (U25749) and a variant from the same species that infects Muscovy ducklings but not goslings, called Muscovy duck parvovirus (DPV; U22967), were first cloned and sequenced in 1995 (Zádori et al., 1995). Vaccines are available and can control infections, but variants arise with some frequency, particularly in China (Ning et al., 2017), and are associated with different syndromes or host ranges, sometimes in association with circoviruses (Li et al., 2018).
Dependoparvovirus is the only genus of Parvovirinae whose members infect reptiles. Serpentine adeno-associated virus 1 was detected as a contaminant of a snake adenovirus 1 isolate (Farkas et al., 2004). Bearded dragon parvovirus, however, has also been detected in deceased bearded dragon specimens where congruent large DNA virus infection could not be confirmed; hence the helper-dependence of this virus is still controversial (Pénzes et al., 2015).
Considerations based on intron length and the phylogeny of Rep amino acid sequences suggest that the monophyletic reptilian viruses are the most basal cluster in the genus (Pénzes et al., 2015).
Species demarcation criteria
Viruses within a species are monophyletic and encode replication initiator proteins (called NS1 or Rep1, 68, or 78) that show >85% amino acid sequence identity.
Related, unclassified viruses
|
Virus name |
Accession number |
Virus abbreviation |
|
murine adeno-associated virus 1 |
MAAV1 |
|
|
murine adeno-associated virus 2 |
MAAV2 |