

Family: Flaviviridae
Genus: Pestivirus
Distinguishing features
Compared to the other viruses in the Flaviviridae, pestiviruses encode two unique gene products, namely Npro and Erns (Tautz et al., 2015). The first protein of the ORF, nonstructural protein Npro, which possesses an autoproteolytic activity and is responsible for its release from the nascent polyprotein (Gottipati et al., 2014, Rumenapf et al., 1998, Stark et al., 1993), is not essential for virus replication in cell culture (Tratschin et al., 1998). One of the three viral envelope glycoproteins, Erns, possesses an intrinsic RNase activity (Krey et al., 2012, Schneider et al., 1993). Both of these unique proteins of the pestiviruses are involved in repression of the host type I IFN response (Schweizer and Peterhans 2001, Meyers et al., 2007, Ruggli et al., 2005, Ruggli et al., 2009, Python et al., 2013, Zurcher et al., 2014, Hilton et al., 2006, La Rocca et al., 2005). Two biotypes of pestiviruses, cytopathogenic (cp) and non-cytopathogenic (noncp) viruses, are distinguished by their ability to cause cytopathic effects in cell culture (Tautz et al., 2015, Becher and Tautz 2011).
A Table with current (MSL38) and previous (MSL37) species names is available at Species names: Flaviviridae.
Virion
Morphology
Virions are 40–60 nm in diameter and spherical in shape (Figure 1. Pestivirus) (Laude 1979). The virion envelope has 10–12 nm ring-like subunits on its surface. The structure and symmetry of the core have not been characterized.
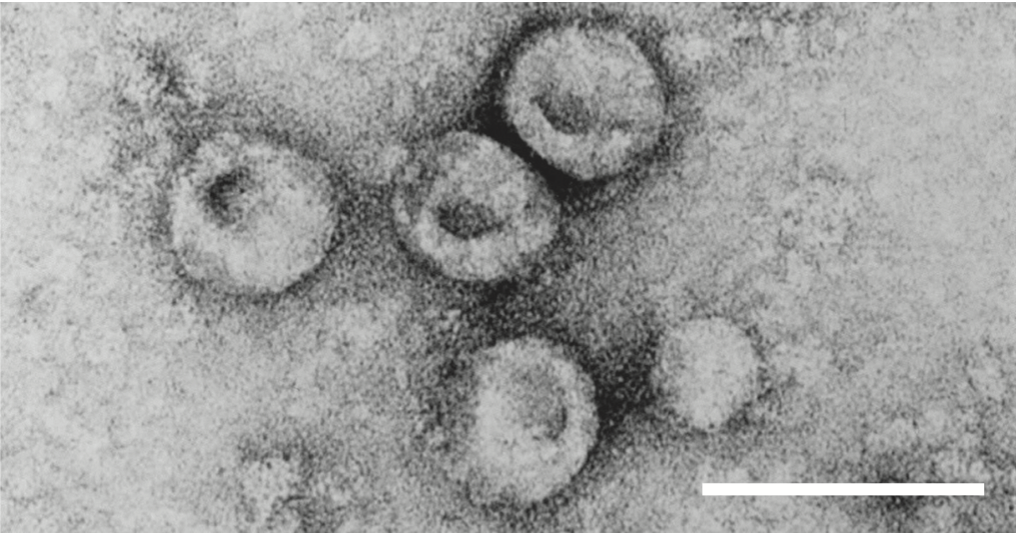 |
| Figure 1. Pestivirus. Negative-contrast electron micrograph of particles of an isolate of bovine viral diarrhea virus 1. The bar represents 100 nm. (From M. König, with permission.) |
Physicochemical and physical properties
Virion Mr has not been determined precisely. Buoyant density in sucrose is 1.10–1.15 g cm−3; S20,W is 140–150S (Laude 1979, Maurer et al., 2004). Virion infectivity is stable over a relatively broad pH range, but unstable at temperatures above 40 °C. Organic solvents and detergents rapidly inactivate these viruses (Depner et al., 1992).
Nucleic acid
The virion RNA is a positive-sense, infectious molecule of ssRNA of 11.3–13.0 kb encoding a single ORF (Becher et al., 2014, Becher et al., 1998, Collett et al., 1988, Meyers et al., 1989). The 5′-NCR contains an IRES and is about 370–385 nt (Poole et al., 1995). The 3′-NCR, of about 185–273 nt, is complex and contains a region with variable sequences and a highly conserved terminal region (Pankraz et al., 2005, Yu et al., 1999). For some cp pestivirus strains, a small and variable segment of host cell or viral nucleic acid is integrated into particular regions (often within NS2 or directly upstream of NS3) of the viral genome, sometimes accompanied by viral gene duplications or deletions (Tautz et al., 2015, Becher and Tautz 2011). Other cp pestiviruses contain only viral gene duplications involving all or part of the Npro and NS3 to NS4B protein-coding regions, resulting in genomic RNA of up to about 16.5 kb. In all cases, the single large ORF is maintained. Finally, cp viruses may also arise by deletion of large portions of their genomes. Such defective genomes can be rescued by co-infecting intact helper viruses (Tautz et al., 2015, Becher and Tautz 2011, Abbas et al., 2013).
Proteins
Virions contain four structural proteins: a basic nucleocapsid core protein, C (14 kDa) and three envelope glycoproteins, Erns (gp44/48), E1 (gp33) and E2 (gp55). All three glycoproteins exist as intermolecular disulfide-linked complexes: Erns homodimers, E1-E2 heterodimers and E2 homodimers (Rumenapf et al., 1991, Thiel et al., 1991, Weiland et al., 1992, Weiland et al., 1990). The Erns protein possesses an intrinsic RNase activity. Pestiviruses encode eight nonstructural (NS) proteins among which Npro (23 kDa), p7 (7 kDa) and NS2 (40 kDa) are not necessary for RNA replication (Behrens et al., 1998, Tautz et al., 1999). Npro is a proteinase that auto-catalytically releases itself from the nascent polyprotein. Nonstructural protein p7 is presumed to have a role in virus maturation (Harada et al., 2000, Elbers et al., 1996). NS2-3 (120 kDa) is a multifunctional protein of which the N-terminal 40% (NS2) is hydrophobic and contains a zinc finger motif that binds divalent metal ions (De Moerlooze et al., 1990, Lackner et al., 2006). NS2 is a cysteine protease that is responsible for processing of NS2-3 to give rise to NS2 and NS3 (Lackner et al., 2006). NS3 (80 kDa) acts as both a serine protease involved in polyprotein processing and an RNA helicase/NTPase involved in RNA replication (Tautz et al., 1997, Tautz et al., 2000, Warrener and Collett 1995, Wiskerchen and Collett 1991). NS2-3 is found after infection with all pestiviruses. In cells infected with cp pestiviruses, large amounts of NS3 can be detected. For noncp bovine viral diarrhea virus (BVDV), noncp Border disease virus (BDV) and classical swine fever virus (CSFV) strains, efficient NS2-3 cleavage is limited to the first eight hours of infection and at later time points the cleavage products NS2 and NS3 are difficult to detect (Lackner et al., 2004). The NS4A (7 kDa) protein acts as a cofactor to the NS3 protease activity (Tautz et al., 2000). The role of NS4B (33 kDa) is unknown. NS5A (58 kDa) represents a phosphorylated protein and also plays still to be further characterized roles in RNA replication and virion morphogenesis (Chen et al., 2012, Isken et al., 2014, Tellinghuisen et al., 2006, Xiao et al., 2009). NS5B (75 kDa) possesses RdRP activity (Zhong et al., 1998, Steffens et al., 1999, Choi et al., 2006).
Lipids
The viruses are enveloped, but no reports have described the lipid composition.
Carbohydrates
All virus envelope glycoproteins contain N-linked glycans (Thiel et al., 1991).
Genome organization and replication
The genomic RNA contains a single large ORF encoding a polyprotein of about 3,900 aa that is preceded by a 5′-NCR of 370–385 nt and followed by a 3′-NCR of 185–273 nt. The gene order is 5′-Npro-C-Erns-E1-E2-p7-NS2-3(NS2-NS3)-NS4A-NS4B-NS5A-NS5B-3′ (Figure 2. Pestivirus) (Tautz et al., 2015, Abbas et al., 2013).
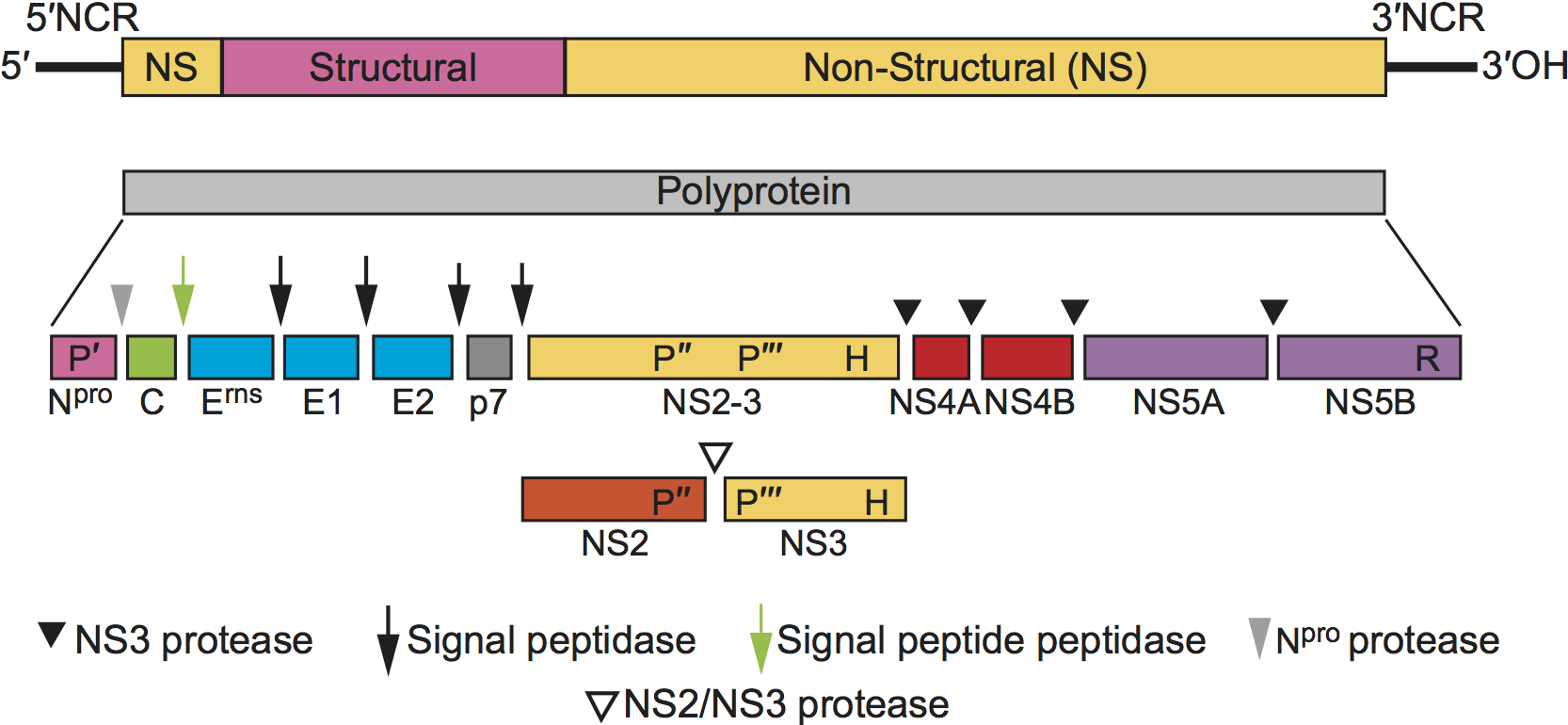 |
| Figure 2. Pestivirus. Pestivirus genome organization (not to scale) and polyprotein processing. The RNA is 11.3–13.0 kb, comprising a 5′-NCR of 370–385 nt, the single ORF of about 11.7 kb and the 3′-NCR of 185–273 nt. Virus nonstructural proteins are indicated as NS. The symbols P′, P″, P′″, H and R indicate the localization of the Npro protease, the NS2 protease, the NS3 protease, the NS3 RNA helicase and the NS5B RdRP, respectively. The proteases and proteolytic steps involved in the generation of individual proteins are indicated. In noncp BVDV viruses, NS2-3 cleavage is detectable early after infection whereas in cp BVDV viruses both NS2-3 and NS3 are produced continuously. |
Pestivirus replication is initiated by receptor-mediated endocytosis involving more than one cell surface molecule and the viral glycoproteins Erns and E2. CD46 has been shown to function as a cellular receptor for BVDV but is not by itself sufficient to mediate infection (Maurer et al., 2004, Krey et al., 2006a, Krey et al., 2006b, Krey et al., 2005). After endocytosis and uncoating, the genome RNA serves as mRNA; there are no subgenomic mRNA molecules. Translation initiation occurs by a cap-independent internal initiation mechanism involving a type IV IRES within the 5′-NCR of the RNA (Lozano and Martinez-Salas 2015). Polyprotein processing occurs co- and post-translationally by both cellular and viral proteases (Tautz et al., 2015). Nonstructural protein Npro, the first protein of the ORF, auto-proteolytically removes itself from the nascent polyprotein by cleavage at the Npro/C site. Downstream cleavages that produce structural proteins C, Erns, E1 and E2 as well as p7 are mediated by cellular signal peptide peptidase and signal peptidase(s) (Elbers et al., 1996, Bintintan and Meyers 2010, Heimann et al., 2006, Rumenapf et al., 1993). Glycoprotein translocation to the endoplasmic reticulum occurs by an internal signal sequence, within the C-terminal region of the C protein. Cleavage between E2 and p7 is not complete, leading to two intracellular forms of E2 with different C-termini (Elbers et al., 1996). Depending on the pestivirus biotype, NS2-3 either remains mostly intact or is found at reduced levels together with high amounts of its N- and C-terminal products NS2 and NS3 (Lackner et al., 2004). The increased generation of NS3 in cp pestiviruses is in most cases due to gene insertion, deletion, duplications or rearrangements (Tautz et al., 2015, Becher and Tautz 2011). The NS3/NS2-3 serine protease activity is responsible for all processing events downstream of NS3. NS4A facilitates cleavages by the NS3 protease of sites 4B/5A and 5A/5B (Tautz et al., 2015).
RNA replication probably occurs in association with intracytoplasmic membranes, presumably in a replication complex composed of viral RNA and viral nonstructural proteins. Nonstructural proteins NS3, 4A, 4B, 5A and 5B are necessary for RNA replication; only NS5A can be provided in trans (Grassmann et al., 2001). Replicative forms of viral RNA have been detected (Gong et al., 1996). The ratio of positive- to negative-sense RNA in cells 12 hours post-infection is about 10. RNA synthesis is resistant to actinomycin D. Virus maturation and release is poorly understood. Budding of virions occurs at ER membranes. Pestivirus particles have been shown in intracellular vesicles and the Golgi complex and during exocytosis (Schmeiser et al., 2014). Considerable amounts of infectious virus remain cell-associated. Host cell RNA and protein synthesis continues throughout infection.
Biology
Pestiviruses infect pigs and ruminants, including cattle, sheep, goats and wild ruminants (Becher et al., 1997, Vilcek and Nettleton 2006). Moreover, pestivirus sequences have been detected in samples from bats and rats by next generation sequencing, but infectious pestiviruses have not yet been isolated from these host species (Firth et al., 2014, Postel et al., 2015, Harasawa et al., 2000, Schirrmeier et al., 2004, Vilcek et al., 2005, Kirkland et al., 2007, Wu et al., 2012, Hause et al., 2015, Postel et al., 2016). There are no invertebrate hosts. Transmission occurs by direct and indirect contact (e.g., nasal or urine secretion, faeces, contaminated food, etc.) and transplacentally. Infections may be subclinical or produce a range of clinical conditions including acute diarrhea, acute hemorrhagic syndrome, acute fatal disease, and a wasting disease. Transplacental infection can result in foetal death, congenital abnormalities, or lifelong persistent infection (Moennig and Plagemann 1992). Fatal mucosal disease can occur in cattle persistently infected with noncp viruses when a cp virus is generated by mutation or introduced by superinfection (Meyers and Thiel 1996). Pestivirus infections of livestock are economically important worldwide (Moennig and Becher 2015, Houe 2003).
Experimental infection models have not been established for bovine or ovine pestiviruses outside their natural mammalian hosts; CSFV can be adapted to propagate in rabbits (Tautz et al., 2015). Cells derived from natural host species (bovine, porcine, ovine) support virus replication. Most virus isolates are noncp and can establish persistent infections in cell culture. Infectious noncp BVDV is often present in bovine serum products used for cell culture (Buttner et al., 1997). Cp pestiviruses induce extensive cytopathology and apoptosis, and form virus plaques under appropriate conditions (Hilbe et al., 2013, Birk et al., 2008, Grummer et al., 2002, Jordan et al., 2002). No hemagglutinating activity has been found associated with pestiviruses.
Antigenicity
Pestiviruses are antigenically related and cross-reactive epitopes have been documented for all species investigated (Tautz et al., 2015, Moennig et al., 1987). Separate antigenic determinants defined by monoclonal antibodies have also been identified. Antigenic variation is particularly pronounced among isolates of BVDV and BDV (Becher et al., 2003, Postel et al., 2015). The N-terminal portion of E2 contains an antigenically hypervariable region (van Rijn et al., 1994, van Rijn et al., 1992). Monoclonal antibody binding patterns are generally consistent with the genetic relatedness of viruses.
Infected animals mount potent antibody responses to two structural glycoproteins (Erns, E2) and to the NS2-3/NS3 protein, while antibody responses to other virus-encoded polypeptides are weak or not detectable. Erns and E2 are able to induce protection independently and monoclonal antibodies reactive with these proteins can neutralize virus infectivity (Weiland et al., 1992, Weiland et al., 1990, Hulst et al., 1993, van Zijl et al., 1991, Reimann et al., 2004).
Species demarcation criteria
Species demarcation criteria in the genus include:
- Nucleotide and deduced amino acid sequence relatedness.
- Antigenic relatedness.
- Host of origin.
Pestiviruses have been assigned to 19 different species based on phylogenetic analysis of conserved amino acid sequences in the regions 189–418, 1,547–2,321, 2,397–2,688 and 3,312– 3,837 (numbered according to the first amino acid of the polyprotein if BVDV-1 Sd-1, M96751) (Figure 3.Pestivirus) (Firth et al., 2014, Postel et al., 2015, Hause et al., 2015, Postel et al., 2016, Smith et al., 2017, Postel et al., 2021, ). Convalescent animal sera generated against members of a given species (e.g., Pestivirus bovis) generally show a several-fold higher neutralization titre against viruses of the same species than against viruses from the other species (Becher et al., 2003, Postel et al., 2015 ). Finally, differences in host of origin and disease can assist in species demarcation.
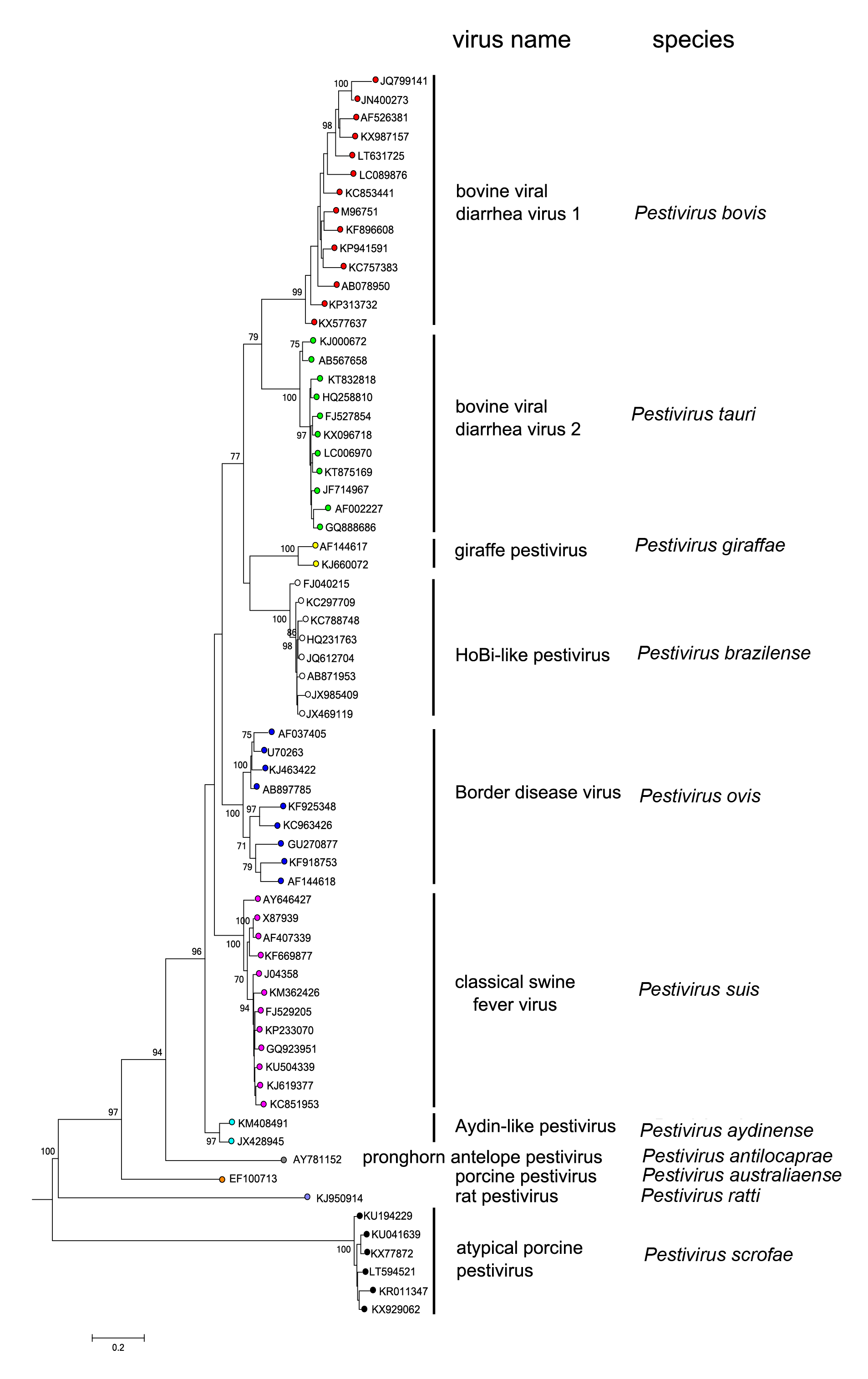 |
| Figure 3. Pestivirus. Phylogenetic tree of pestivirus amino acid positions 3,312–3,899. Maximum likelihood distances were calculated using a JTT+G model in MEGA 6 (Tamura et al., 2013) using up to 15 of the most divergent sequences for each species, with elimination of sequences <1 % divergent. Branches supported by >70 % of bootstrap replicates are indicated. Virus names and species assignments are indicated to the right. This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. |
For example, Pestivirus bovis and Pestivirus suis are considered different species because their members have: (i) amino acid sequences that are phylogenetically distinct in three different subgenomic regions (ii) at least 10-fold difference in neutralization titre in cross-neutralization tests with polyclonal immune sera, and (iii) host range, in that under natural conditions members of Pestivirus suis infect only pigs while members of Pestivirus bovis infect ruminants as well as pigs.
Members of the species Pestivirus bovis can be further subdivided into at least seventeen genotypes, while three genotypes (CSFV-1, CSFV-2, and CSFV-3) are recognized among members of Pestivirus suis (Yesilbag et al., 2014, Postel et al., 2012). Genotypes of these Pestivirus species can be further divided into subgroups.
Proposals for additional pestivirus species should be based on the complete genome sequence of at least one virus isolate, while data on antigenic relatedness and host range should also be considered. An incomplete genome sequence of an isolate from a bat (JQ814854) therefore remains unclassified.
Related, unclassified viruses
| Virus name | Accession number | Virus abbreviation |
| bat pestivirus | JQ814854 | BaPV |
Virus names and virus abbreviations are not official ICTV designations.