

Family: Flaviviridae
Genus: Pegivirus
Distinguishing features
Pegiviruses have been detected in a variety of mammalian hosts, with transmission of human pegiviruses occurring by sexual, parenteral and maternal routes. Invertebrate vectors have not been discovered.
Pegiviruses show distant sequence relatedness to other members of the family Flaviviridae, forming a distinct cluster based on phylogenetic analysis of the RdRP (Figure 1. Flaviviridae). In addition to their separate phylogenetic position, they show several differences in genome organization from members of the Hepacivirus, Orthoflavivirus, and Pestivirus genera. Most pegiviruses possess an IRES element that is structurally unrelated to those of hepaciviruses and pestiviruses and they do not encode a protein homologous to the nucleocapsid protein found in members of the other genera (Quan et al., 2013, Muerhoff et al., 1995, Stapleton et al., 2011). Infections with human pegivirus (HPgV) are frequently persistent but, with the exception of an association with non-Hodgkin’s lymphomac (Krajden et al., 2010, Chang et al., 2014), are not associated with the development of any identifiable disease. Pegivirus infections of other mammalian species are persistent and non-pathogenic, apart from the report of Theiler’s disease in horses infected with Theiler’s disease associated virus (Chandriani et al., 2013).
A Table with current (MSL38) and previous (MSL37) species names is available at Species names: Flaviviridae.
Virion
Morphology
Virions of pegiviruses have not been visualized; the lack of an encoded core protein suggests that they may be structurally distinct from other members of the Flaviviridae. The virion size of HPgV was estimated to be 50–100 nm based on sequential filtration through filters of decreasing pore sizes.
Physicochemical and physical properties
The buoyant density of HPgV from human serum on both sucrose and CsCl density centrifugation ranged from 1.05–1.13 g cm−3 (Xiang et al., 1998, Melvin et al., 1998). Treatment of HPgV with detergent did not recover a denser, non-enveloped form of the virion, consistent with the lack of a viral nucleocapsid (Melvin et al., 1998). In the absence of an established cell culture model for pegiviruses, no information is currently available on their stability or inactivation characteristics.
Nucleic acid
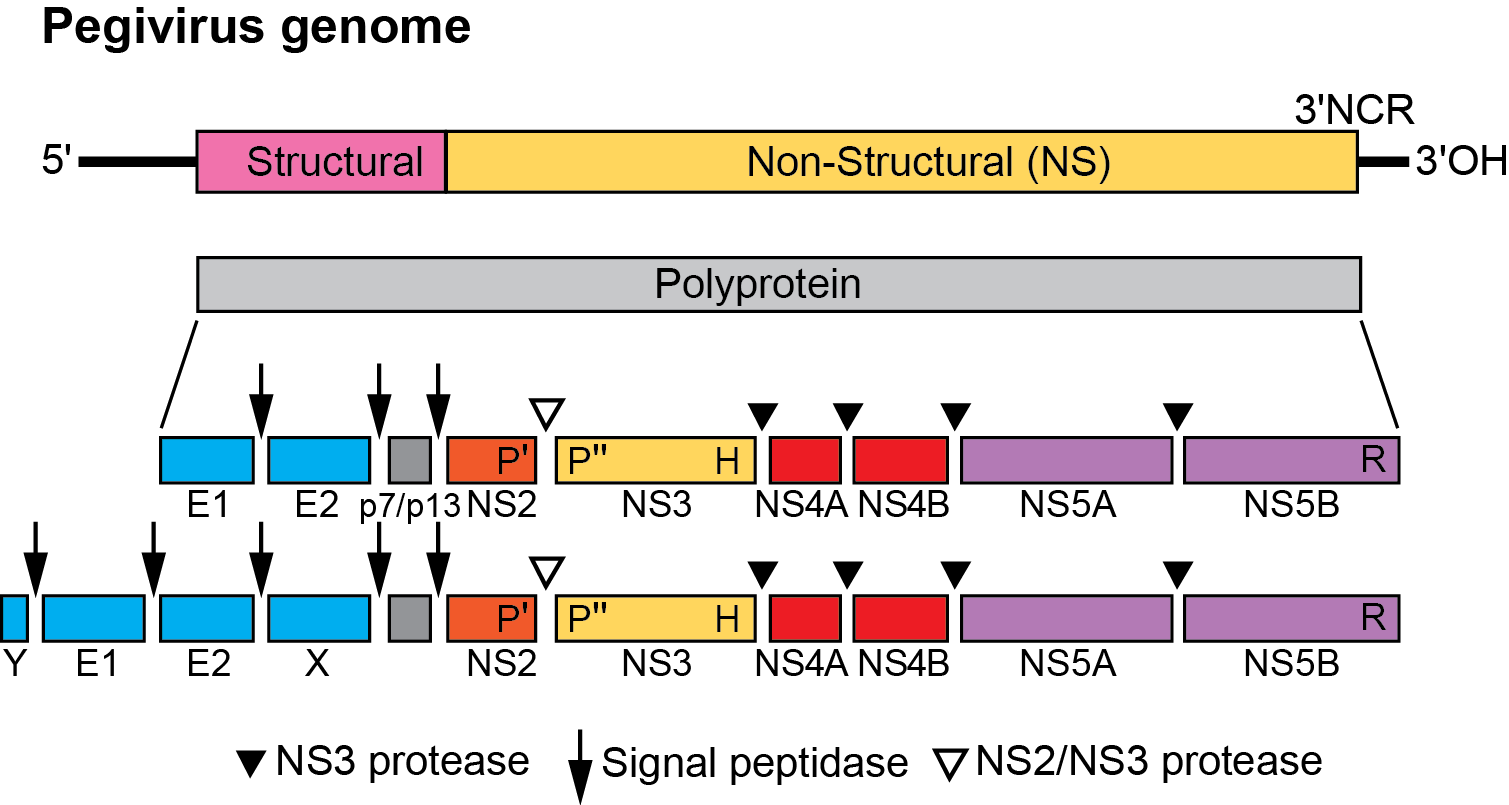
Pegivirus virions contain a single positive-sense, potentially infectious ssRNA ranging from 8.9–11.3 kb (Figure 1. Pegivirus). The 5′-NCR contains an IRES element of between 300-550 nt. No miR-122 binding sites have been identified in 5′-NCR sequences of HPgV or among members of other pegivirus species (Smith et al., 2016). Most pegiviruses possess an IRES broadly similar in structure but not in sequence to the type I IRES elements of picornaviruses (Quan et al., 2013); however, the more divergent human hepegivirus (HHPgV), a member of Pegivirus columbiaense, as well as members of Pegivirus carolliae and Pegivirus neotomae have type IV IRES elements structurally resembling those of hepaciviruses and pestiviruses, but again with almost no sequence identity between them (Kapoor et al., 2015).
Proteins
Functional studies of most pegivirus proteins have not been performed and information on their likely function in replication and virus assembly has largely been inferred from comparison with homologous genes in hepaciviruses. Most pegiviruses lack or possess no obvious homologue of the core protein of hepaciviruses and other members of the Flaviviridae, so how pegivirus virions are assembled remains uncertain. However, typically pegiviruses are predicted to encode shorter, although somewhat variable length, basic proteins containing multiple arginine and leucine amino acids immediately upstream of the signalase site before E1; these may play some role in RNA packaging during virion assembly. E1 and E2 proteins are believed to be envelope glycoproteins, while NS3 and NS5B contain motifs common to helicase and polymerase proteins in viruses of other genera of the Flaviviridae (reviewed in (Stapleton et al., 2011)). The NS3-4A region has been shown to be proteolytically active for processing the nonstructural region of the human pegivirus polyprotein (Belyaev et al., 1998).
Lipids
The virion structure of pegiviruses is unknown, but the presence of predicted hydrophobic transmembrane regions in the E1 and E2 glycoproteins is consistent with the presence of viral envelope, likely derived by budding of pegiviruses from infected cells, analogously to other members of the family Flaviviridae.
Carbohydrates
The E1 and E2 glycoproteins have variable numbers of potential N-linked glycosylation sites, with members of the more divergent species Pegivirus columbiaense, Pegivirus carolliae and Pegivirus neotomae possessing a larger number of sites, a feature more typical of hepaciviruses (Kapoor et al., 2015).
Genome organization and replication
In common with other members of the Flaviviridae, the genome contains a single ORF. Structural proteins are processed by cellular proteases, while the NS3-4A viral protease cleaves the nonstructural region of the polyprotein in the same gene order as hepaciviruses (Figure 1.Pegivirus).
|
|
|
Figure 1.Pegivirus. Genome organization of pegiviruses. Pegivirus genomes range from approximately 8.9–11.3 kb; those with longer genomes code for additional predicted structural proteins, X and Y (lower diagram). The genome encodes a polyprotein that is co- and post-translationally cleaved into individual viral proteins. Structural proteins common to all pegiviruses are the envelope glycoproteins (E1 and E2), and non-structural proteins are NS2–NS5B. No protein homologous to the core protein of other members of the Flaviviridae has been identified in pegiviruses although some possess a predicted, basic protein upstream of E1 of unknown function (Y). Several pegiviruses also have a predicted additional glycoprotein downstream of E2 (X). Cleavage of structural proteins by cellular signal peptidases, of NS2/NS3 by the NS2–NS3 autoprotease and of the remaining NS proteins by the NS3–NS4A protease complex is comparable to hepaciviruses. All pegiviruses possess a long 5ʹ-noncoding region with predicted IRES function; most pegiviruses have a type I picornavirus-like IRES while others have a type IV IRES type structurally related to those of hepaciviruses and pestiviruses. |
Biology
Host range
Pegiviruses can be detected in a wide range of mammalian species (humans, non-human primates, pigs, horses and a range of rodent and bat species). Members of Pegivirus platyrrhini infect New World monkeys (Muerhoff et al., 1995, Bukh and Apgar 1997) and bats, while members of Pegivirus hominis infect humans, chimpanzees (Adams et al., 1998, Birkenmeyer et al., 1998) and Old World monkeys (Sibley et al., 2014, Bailey et al., 2016). Members of Pegivirus caballi (equine pegivirus (Kapoor et al., 2013a) and Pegivirus equi (Theiler’s disease associated virus (Chandriani et al., 2013)) infect horses, members of Pegivirus suis infect pigs (Baechlein et al., 2016) and members of Pegivirus carolliae, Pegivirus neotomae, Pegivirus pteropi, Pegivirus scotophili and Pegivirus sturnirae infect a wide range of bat and rodent species (Kapoor et al., 2013b, Quan et al., 2013, Epstein et al., 2010)(Epstein et al. , 2010; Kapoor et al. , 2013b; Quan et al. , 2013).
Very limited information is available on the potential of pegiviruses to transmit between different host species. However, chimpanzees can be experimentally infected by inoculation with HPgV (Pegivirus hominis) but not by the New World primate virus, GBV-A (Pegivirus platyrrhini) (Bukh et al., 1998) and rhesus macaques can be infected with a baboon isolate of Pegivirus hominis (Bailey et al., 2015).
Transmission
HPgV is transmitted between humans by sexual transmission, exposure to contaminated blood, and from mother to child. Horizontal transmission has neither been confirmed nor refuted (Bhattarai and Stapleton 2012). HPgV viraemia frequencies are higher in injecting drug users and in haemophiliacs with a history of exposure to non-virally inactivated clotting factor concentrates, indicating an efficient parenteral route of transmission. HPgV viraemia frequencies are also higher in people with sexually transmitted diseases and without a history of parenteral exposure (Scallan et al., 1998); human pegivirus infection is also a frequent co-infection with human immunodefiency virus 1 (HIV-1). Among HIV-infected subjects, co-infection with HPgV does not correlate with HIV transmission risk; however, hepatitis C virus (HCV) and HCV-HPgV co-infection are significantly associated with a parenteral mode of HIV acquisition (Bourlet et al., 1999), indicating the likelihood of sexual routes of transmission.
HHPgV (Pegivirus columbiaense) additionally shows evidence for parenteral routes of transmission with infections largely confined to intravenous drug users and recipients of blood or blood products (Kapoor et al., 2015, Coller et al., 2016, Bonsall et al., 2016, Berg et al., 2015); however, other modes of transmission have not been extensively studied.
Geographical distribution
Infection of humans with HPgV occurs worldwide and it is likely that it is ubiquitous in human populations. Prevalence studies in developed countries indicate between 1–4 % of healthy blood donors are viraemic for HPgV and another 5–13 % have anti-E2 antibodies, indicating prior infection. Rates of infection with HPgV in developing countries are higher, with viraemia frequencies in the general population frequently exceeding 10%. Infection frequencies of pegiviruses infecting non-human hosts are incompletely described. Infections with HHPgV appear to be confined to those with parenteral exposure, more reminiscent of HCV. Relatively low frequencies (1%–2%) of viraemia of other pegiviruses have been described in horses for members of Pegivirus caballi and Pegivirus equi (Lu et al., 2016, Lyons et al., 2014, Kapoor et al., 2013a, de Souza et al., 2015) and pigs for members of Pegivirus suis) (Baechlein et al., 2016).
Pathogenicity
Infections with HPgV in humans are considered non-pathogenic, to the extent that viraemic blood donations are not excluded from transfusion. The pathogenicity of pegiviruses infecting other hosts is unknown although experimental infection of New World primates with simian pegiviruses does not induce liver disease. However, it has been reported that infection of horses with members of Pegivirus equi is associated with Theiler’s disease in horses (Chandriani et al., 2013).
Cell tropism
Pegiviruses infecting humans or new world primates cannot be readily detected in the liver of infected hosts, whereas they are present at higher viral loads in circulating lymphocytes, including T and B lymphocytes (Kobayashi et al., 1999, Tucker et al., 2000). However, based on autopsy studies in humans (Tucker et al., 2000, Radkowski et al., 1999), and the animal model of nonhuman pegivirus infection in rhesus macaques, pegivirus replication occurs primarily in the bone marrow (Bailey et al., 2015). The tissue or cellular tropism of pegiviruses infecting other hosts is unknown. Like the hepaciviruses, pegiviruses differ from members of the genera Orthoflavivirus and Pestivirus by their limited ability to be propagated in cell culture (Chivero and Stapleton 2015).
Antigenicity
Pegivirus antigenicity is poorly characterized in the absence of in vitro neutralization assays or experimental animal models. Antibody to the E2 glycoprotein of HPgV can be detected in humans and is associated with clearance of viraemia (Feucht et al., 1997, Tacke et al., 1997). These E2 antibodies reduce the rate of re-infection following liver transplantation (Tillmann et al., 1998). Recent data show the immune modulating effects of E2 protein on T cell activation and NK cell signalling, which may contribute to the absence of serological reactivity to other HPgV proteins (Chivero et al., 2015).
Species demarcation criteria
Species in the Pegivirus genus are now classified based on their genetic divergence (Smith et al., 2016) rather than their host range (Stapleton et al., 2011). Assignment thresholds are based on amino acid sequence divergence in conserved regions of NS3 and NS5B; pegiviruses showing greater than 0.31 amino acid p-distances in a conserved region of NS3 (positions 888–1635 as numbered in the HPgV reference sequence U22303), and greater than 0.31–0.36 in the NS5B region (amino acid positions 2398–2916) are considered to be separate species (Smith et al., 2016). In general, members of different pegivirus species infect different hosts with the notable exception of Pegivirus platyrrhini, into which are assigned pegiviruses infecting New world primates (GB virus-A) and African bats.
Isolates of the species Pegivirus hominis are also known as hepatitis G virus (HGV) (Linnen et al., 1996, Simons et al., 1995a), although more recently the name human pegivirus (HPgV) has been proposed and adopted as there is now no evidence that infections are associated with hepatitis (Stapleton et al., 2011), nor did this virus infect the surgeon, GB, from whom infective material was passaged in primates and the virus first described. A second, more divergent group of pegiviruses, termed human hepegivirus (HHPgV) or HPgV-2 (Kapoor et al., 2015, Berg et al., 2015) has been assigned to Pegivirus columbiaense (Smith et al., 2016).