

Family: Iridoviridae
V. Gregory Chinchar, Paul Hick, Jie Huang, Ikbal Agah Ince, James K Jancovich, Rachel Marschang, Qiwei Qin, Kuttichantran Subramaniam, Thomas B Waltzek, Richard Whittington, Trevor Williams and Qi-Ya Zhang
The citation for this ICTV Report chapter is the summary published as Chinchar et al., (2017):
ICTV Virus Taxonomy Profile: Iridoviridae, Journal of General Virology, 98, 890–891.
Corresponding author: V Gregory Chinchar ([email protected])
Edited by: Balázs Harrach and Andrew J. Davison
Posted: May 2017, updated May 2018 & February 2020
PDF: ICTV_Iridoviridae.pdf
Summary
Members of the family Iridoviridae comprise a collection of large icosahedral, double-stranded DNA (dsDNA)-containing viruses that are classified into two subfamilies: Alphairidovirinae and Betairidovirinae (Table 1 Iridoviridae). The former is comprised of three genera (Ranavirus, Megalocytivirus and Lymphocystivirus) whose members infect primarily ectothermic vertebrates such as bony fish, amphibians and reptiles, whereas the latter contains three genera (Iridovirus, Chloriridovirus, and Decapodiridovirus) whose members infect mainly invertebrates such as insects and crustaceans. Viral macromolecular synthesis involves both nuclear and cytoplasmic compartments and virion assembly takes place in the cytoplasm within morphologically distinct viral assembly sites. Mature, non-enveloped, but otherwise infectious, virions may remain within the cytoplasm from which they are released by cell lysis, or virions may acquire an envelope by budding from the plasma membrane. To avoid confusion between members of the genus Iridovirus and members of the family, members of the family will be referred to as iridovirids, rather than iridoviruses.
Table 1 Iridoviridae. Characteristics of members of the family Iridoviridae.
| Characteristic | Description |
| Typical member | frog virus 3 (AY548484), species Ranavirus rana1 |
| Virion | Typically 150–200 nm (non-enveloped); the principal component of the capsid is the 48 kDa major capsid protein |
| Genome | Linear, double-stranded DNA, 103–220 kbp, encoding 92–211 putative proteins ; the genome is circularly permuted and terminally redundant |
| Replication | First stage DNA synthesis and early transcription take place in the nucleus; subsequently DNA concatemer formation and late transcription occur in the cytoplasm; virion morphogenesis takes place in cytoplasmic assembly sites |
| Translation | Directly from capped, non-polyadenylated mRNAs |
| Host Range | Subfamily Alphairidovirinae, members infect amphibians, fish, reptiles; subfamily Betairidovirinae, members infect mainly insects and crustaceans. |
| Taxonomy | Realm Varidnaviria, kingdom Bamfordvirae, phylum Nucleocytoviricota, class Megaviricetes, order Pimascovirales: seven genera divided between two subfamilies with 22 species |
Virion
Morphology
Virions consist, of a central DNA/protein core surrounded in turn by an internal limiting membrane, a viral capsid, and, in the case of those particles that bud from the plasma membrane, an outer viral envelope (Figure 1. Iridoviridae). SDS-PAGE analysis indicates the presence of approximately 30 virion-associated proteins, while proteomic analyses suggest higher numbers (Wong et al., 2011, Ince et al., 2015). Although some virion-associated proteins such as the major capsid protein, comprise structural elements of the particle, others serve catalytic or regulatory roles, e.g., the virion-associated transactivator of immediate-early transcription. Viral capsids display icosahedral symmetry (Figure 2A. Iridoviridae) and are usually 120–200 nm in diameter, but may be up to 350 nm (e.g. genus Lymphocystivirus). The virion core is electron-dense and consists of a DNA-protein filament enclosed within an icosahedral capsid (Figure 1. Iridoviridae).
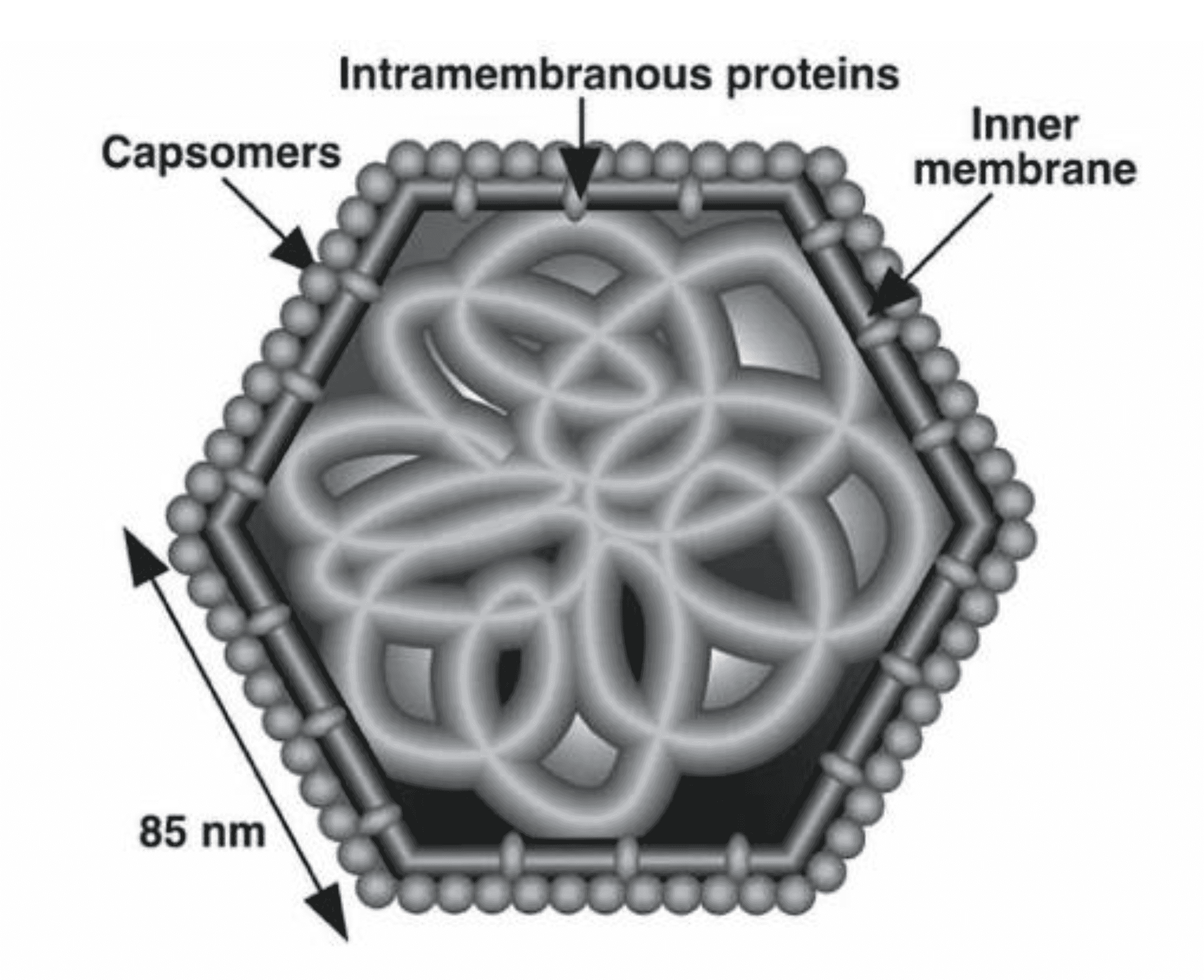 |
| Figure 1. Iridoviridae. Schematic diagram of a cross-section of an iridovirus particle, showing capsomers, the internal lipid bilayer containing transmembrane proteins and the filamentous nucleoprotein core (from (Darcy-Tripier et al., 1984) with permission). |
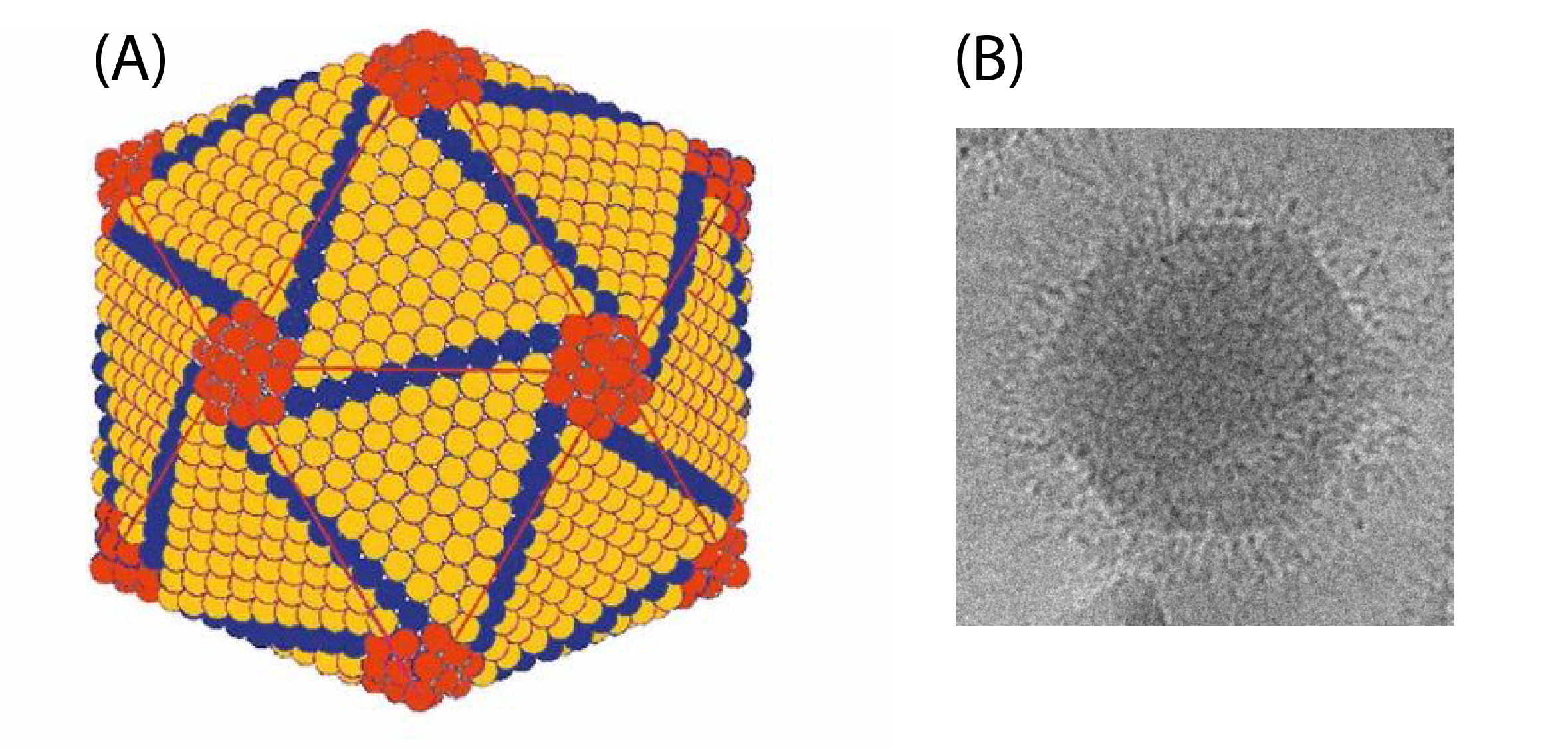 |
| Figure 2. Iridoviridae. A: Outer shell of invertebrate iridescent virus 2 (IIV2). Shown are trisymmetrons (orange) comprising the 20 icosahedral faces and pentrasymmetrons (red) located at the 12 vertices, and disymmetrons (blue) located at the edges of the icosahedral faces (Adapted from (Wrigley 1969) with permission). B: Cryo-transmission electron micrograph of IIV6 displaying a fringe of hair-like fibers surrounding the viral particle (figure provided by IA Ince and J van Lent). |
Each virion is formed of 12 pentasymmetrons and 20 trisymmetrons arranged in an icosahedral, quasi-equivalent symmetry with a triangulation number T=147 (h=7, k=7) (Yan et al., 2009). Both types of structure predominantly comprise hexavalent capsomers, a total of 1,460 per virion, that are composed of the major capsid protein (MCP). Each hexavalent capsomer is formed by a non-covalent MCP trimer on the outer surface and a second MCP trimer linked by disulfide bonds on the inner surface. In addition, each pentasymmetron comprises 30 hexavalent (trimeric) capsomers and a single pentavalent capsomer at its center, at the vertex of each pentasymmetron, for a total of 12 in each virion.
Several proteins have been identified in the capsid shell and in association with the internal lipid membrane (Yan et al., 2009). These have been named zip monomers, zip dimers, finger proteins and anchor proteins. The zip dimers appear as two halves of a clasp connecting the trimer capsomers along the edges of adjacent trisymmetrons, whereas zip monomers appear to be involved in linking trisymmetron capsomers with those of neighboring pentasymmetrons. Three sets of nine inward-pointing finger proteins bind the capsomers along the edges of each trisymmetron. Finally, an anchor protein connects each pentasymmetron with the lipid membrane at a distance of two capsomers from the pentavalent vertex. Other transmembrane proteins are present, but only the anchor protein can be visualized in image reconstruction studies due to its invariant position with respect to the symmetry of the particle. Size estimates using the MCP as reference suggest molecular masses of 11.9, 19.7 and 32.4 kDa for the zip, finger and anchor proteins, respectively.
Among members of the Iridovirus, Chloriridovirus and Lymphocystivirus genera, the outer surface of the capsid is covered by flexible fibrils (Figure 2B. Iridoviridae). Conventional EM studies suggest that these fibrils are often rather short (ca. 2.5 nm in length) and may have terminal knobs, but in IIV6 a single fibril 2 nm wide and about 35 nm long extends outwards from the center of each trimeric capsomer. The role of the fibrils remains unknown. Fibrils have not been reported among ranaviruses or megalocytiviruses.
Iridovirids may acquire an envelope by budding through the host cell membrane. The envelope increases the specific infectivity of virions, but is not required for infectivity as naked particles are also infectious.
Physicochemical and physical properties
The Mol wt of virions is 1.05–2.75×109 with a sedimentation coefficient (S20,w) of 2020–4460S and a density of 1.26–1.60 g cm−3. Virions are stable in water at 4°C for extended periods. Iridovirids are inactivated by pH <3.0 and >11.0 and by exposure to UV-irradiation on the order of 103 μWs/cm2. Virions are inactivated within 30 min at >55°C. Some ranaviruses remain infectious after desiccation, e.g., Bohle iridovirus (BIV) survives desiccation at temperatures up to 42°C for up to 6 weeks, whereas others are sensitive to drying.
Nucleic acid
Virions contain a single linear double-stranded DNA molecule of 140–303 kbp, a value that includes both unique and terminally redundant sequences. However, when considering only the unique portion, genomes range in size from 103 to 220 kbp, depending upon the specific viral species (Table 2. Iridoviridae). DNA comprises 12–16% of the particle weight, and the G+C content ranges from about 27 to 55%. All viruses within the family possess genomes that are circularly permuted and terminally redundant. The DNA of vertebrate iridoviruses (subfamily Alphairidovirinae) is, with the exception of Singapore grouper iridovirus (SGIV), highly methylated, whereas little to no methylation is found within the genomes of the invertebrate iridoviruses (subfamily Betairidovirinae). The complete genomic sequence is known for over 40 members of the family with representative sequence information available for every genus (Table 2. Iridoviridae). Although naked genomic DNA is not infectious, non-genetic reactivation of viral DNA can be achieved in the presence of viral structural proteins.
Table 2. Iridoviridae. Iridovirus genome content.
| Subfamily Alphairidovirinae | ||||||
| Genus | Species | Isolate | Genome (bp) | ORFs | %GC | Accession number |
| Lymphocystivirus | Lymphocystis disease virus 1 | lymphocystis disease virus 1 (LCDV1) | 102,653 | 108 | 29 | L63545 |
| Lymphocystis disease virus 2 | lymphocystis disease virus-China (LCDV-C) | 186,250 | 176 | 27 | AY380826 | |
| Lymphocystis disease virus 3 | lymphocystis disease virus-Sparus aurata (LCDV-Sa) | 208,501 | 183 | 33 | KX643370 | |
| Ranavirus | Frog virus 3 | frog virus 3 (FV3) | 105,903 | 97 | 55 | AY548484 |
| Ambystoma tigrinum virus | Ambystoma tigrinum virus (ATV) | 106,332 | 92 | 54 | AY150217 | |
| Epizootic haematopoietic necrosis virus | epizootic hematopoietic necrosis virus (EHNV) | 127,011 | 100 | 54 | FJ433873 | |
| Common midwife toad virus | common midwife toad virus (CMTV) | 106,878 | 104 | 55 | JQ231222 | |
| Santee-Cooper ranavirus | largemouth bass virus (LMBV) | NDa | ND | ND | DQ159940 | |
| Singapore grouper iridovirus | Singapore grouper iridovirus (SGIV) | 140,131 | 139 | 49 | AY521625 | |
| Megalocytivirus | Infectious spleen and kidney necrosis virus | infectious spleen and kidney necrosis virus (ISKNV) | 111,362 | 117 | 55 | AF371960 |
| Scale drop disease virus | scale drop disease virus (SDDV) | 124,244 | 129 | 37 | KR139659 | |
| Subfamily Betairidovirinae | ||||||
| Genus | Species | Isolate | Genome (bp) | ORFs | %GC | Accession number |
| Iridovirus | Invertebrate iridescent virus 6 | invertebrate iridescent virus 6 (IIV6) | 212,482 | 211 | 29 | AF303741 |
| Invertebrate iridescent virus 31 | invertebrate iridescent virus 31 (IIV31) | 220,222 | 203 | 35 | HF920637 | |
| Chloriridovirus | Invertebrate iridescent virus 3 | invertebrate iridescent virus 3 (IIV3) | 191,132 | 126 | 48 | DQ643392 |
| Invertebrate iridescent virus 9 | invertebrate iridescent virus 9 (IIV9) | 206,791 | 191 | 31 | GQ918152 | |
| Invertebrate iridescent virus 22 | invertebrate iridescent virus 22 (IIV22) | 197,693 | 167 | 29 | HF920633 | |
| Invertebrate iridescent virus 25 | invertebrate iridescent virus 25 (IIV25) | 204,815 | 177 | 30 | HF920635 | |
| Anopheles minimus iridovirus | Anopheles minimus iridovirus (AMIV) | 163,023 | 148 | ND | KF938901 | |
| Decapod-iridovirus | Decapod iridescent virus 1 | Shrimp hemocyte iridescent virus (SHIV) - 20141215 | 165,809 | 170 | 35 | MF599468 |
a ND: not determined. The genome sequences of largemouth bass virus, doctor fish virus and guppy virus 6 are unpublished, but the nucleotide sequences of their 26 core protein genes have been determined and aligned with those of other members of the family; all three isolates are placed in the species Santee-Cooper ranavirus. The accession number listed is for the major capsid protein gene of the largemouth bass virus isolate.
Proteins
Nucleotide sequence analysis has identified more than 100 ORFs among various members of the family (Tables 2 and 3. Iridoviridae, Figure 3. Iridoviridae) of which 26 are common to all iridovirids (Eaton et al., 2007). As discussed below, viral proteins can be ordered into one of three, somewhat overlapping, categories: catalytic, structural, and virulence-related.
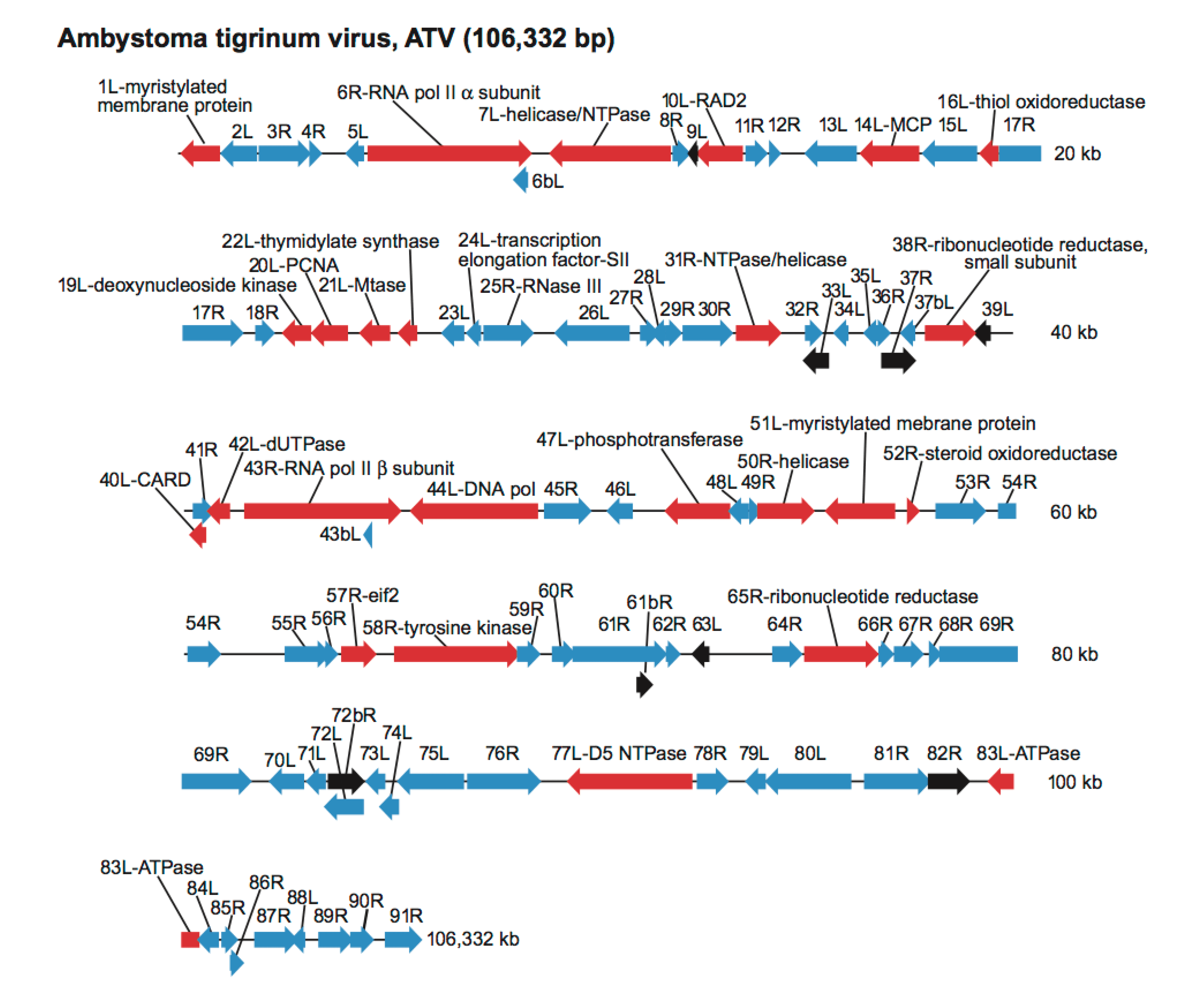 |
| Figure 3. .Iridoviridae. Genetic organization of a representative member of the family Iridoviridae, Ambystoma tigrinum virus (ATV), species Ambystoma tigrinum virus, genus Ranavirus. Arrows represent viral ORFs with their size, position and orientation shown. ORFs of known function are colored red and their putative proteins identified; ORFs with known homology to tiger frog virus (TFV), an isolate of Frog virus 3, but of unknown function are shaded blue; and those of unknown function or with no homology to TFV are indicated in black. |
Catalytic proteins comprise the majority of the 26 conserved proteins and include the virus-encoded DNA polymerase, the two largest subunits of the viral homolog of RNA polymerase II (vPOL-II), and the small subunit of ribonucleotide reductase (Table 3. Iridoviridae). In most cases their assigned function is putative and based on homology with other proteins in the database. However, in some cases, function has been confirmed experimentally, e.g., knock down of vPOL-IIα using an antisense morpholino oligonucleotide results in a marked reduction in late viral gene expression and supports its role as a viral transcriptase.
Iridovirid particles are complex assemblies of both structural and virion-associated proteins. Aside from structural elements such as the MCP and the aforementioned zip, finger, and anchor proteins, virions contain an additional 30 (or more) proteins ranging in mass from 5 to 250 kDa. The major capsid protein (MCP, 48–55 kDa) is highly conserved among all iridovirids and comprises 40% of the total virion protein mass. The amino acid (aa) sequence of the MCP has been used in virus identification and in phylogenetic analyses.
In addition to structural proteins that contribute to the shape and integrity of the virion, there are a number of virion-associated proteins that play specific roles in viral replication. For example, virions contain a putative transcriptional transactivator that directs the expression of immediate-early viral genes. Furthermore, at least six DNA-associated polypeptides have been identified within the core of IIV6 including a major species of 12.5 kDa. Likewise, a number of virion-associated enzymatic activities have been detected, including a protein kinase, nucleotide phosphohydrolase, a ss/dsRNA-specific ribonuclease, pH 5 and pH 7.5 deoxyribonucleases, a protein phosphatase and a protein that triggers the shutdown of host macromolecular synthesis. In addition to these polypeptides, various other putative catalytic proteins have been identified by BLAST analysis some of which are encoded by all family members (Table 3. Iridoviridae; Figure 3. Iridoviridae).
Lastly several putative virulence or immune evasion proteins that enhance viral replication in restrictive environments (e.g., quiescent cells) or block host immune responses have been detected (Grayfer et al., 2015). Among the former are viral homologs of dUTPase, thymidine kinase, and ribonucleotide reductase that maintain deoxyribonucleotide pools during viral infection, whereas among the latter, vCARD, vIF-2α, vRNase III, and β-steroid dehydrogenase may function to block innate and acquired immunity (Grayfer et al., 2015). For example, vIF-2α, the viral homolog of eukaryotic translational initiation factor eIF-2α, blocks translational shut-off mediated by protein kinase R and maintains viral protein synthesis during infection (Rothenburg et al., 2011).
Table 3. Iridoviridae. Core genes of members of the family Iridoviridaea
| ORFb | Putative function | Categoryc | ORFb | Putative function | Categoryc |
| 1R | Putative replication factor and/or DNA binding/packaging protein | C | 57R | Serine/threonine protein kinase | C |
| 2L | Myristylated membrane protein | S | 60R | DNA polymerase family B exonuclease | C |
| 8R | DNA-dependent, RNA polymerase II, α subunit | C | 62L | DNA-dependent, RNA polymerase II, β subunit | C |
| 9L | NTPase/helicase | C | 67L | Ribonucleotide reductase, small subunit (RRα) | C,V |
| 12L | Unknown function | UNK | 80L | RNase III | C,V |
| 15R | AAA-ATPase, similar to poxvirus A32, required for DNA packaging | C | 81R | Transcription elongation factor TFIIS | C |
| 19R | Serine/threonine protein kinase | C | 84R | Proliferating cell nuclear antigen (PCNA) | C |
| 21L | Helicase family protein | C | 85R | Deoxynucleoside kinase | C |
| 22R | D5 family NTPase involved in DNA replication | C | 88R | Erv/Alr family of thiol oxidoreductases | C,V |
| 27R | Putative tyrosine kinase/LPS modifying enzyme | C,V | 90R | Major capsid protein (MCP) | S |
| 37R | NIF-NLI interacting factor | V | 91R | Immediate early protein ICP-46
| UNK |
| 41R | Vaccinia virus early transcription factor | C | 94L | Uvr/REP helicase | C |
| 53R | Myristylated membrane protein | S | 95R | Putative RAD2-type nuclease | C |
a Two iridovirids contain only 25 of the 26 core genes: SHIV lacks the gene for the small subunit of ribonucleotide reductase; ECIV lacks the gene for deoxynucleotide kinase. In addition to the 26 iridovirid core genes, 27 ranavirus specific genes and 13 amphibian-like ranavirus genes have been identified (Eaton et al., 2007, Jancovich et al., 2010). Of the 27 ranavirus-specific genes 24 are of unknown function, whereas the three that are known encode a CARD-containing protein (64R), dUTPase (63R) and neurofilament triplet H1-like protein (32R). Likewise only two of the 13 amphibian ranavirus-like genes (26R, vIF-2α; 82R, immediate early 18 kDa protein) have putative homologs.
b ORF designations are based on their positions within the genome of frog virus 3 (AY548484).
c The letters reflect the known or putative function of the indicated proteins: C, catalytic; S, structural, V, virulence; UNK, unknown. In addition, several catalytic proteins are likely found within virions as virion-associated proteins.
Lipids
Non-enveloped particles contain 5–17% lipid, predominantly as phospholipid. Virions possess an internal lipid membrane that lies between the DNA core and the viral capsid, and, in budded virions, an outer viral envelope is acquired from the plasma membrane. The origin of the internal lipid membrane is unclear but, based on evidence from other nuclear cytoplasmic large DNA viruses (NCLDV) such as vaccinia virus, African swine fever virus, and Paramecium bursaria chlorella virus, it is likely derived from breakdown of the host endoplasmic reticulum (Romero-Brey and Bartenschlager 2016, Milrot et al., 2016). By analogy to other NCLDVs the internal membrane likely plays a key role in virion assembly by serving as a scaffold on which capsid protein deposition takes place.
Carbohydrates
Carbohydrate-containing proteins (glycoproteins) have been detected within lymphocystis disease virus and are likely present within other members of the family (Garcia-Rosado et al., 2004).
Genome organization and replication
Iridovirid genomes consist of a unique component and a variable amount of terminal redundancy. The unique component ranges from 103 to 220 kbp, whereas the actual genome content is 5–50% larger due to the presence of terminal redundancy and varies depending upon species. With the exception of LCDV1 (103 kbp), genomes for members of the Lymphocystivirus, Iridovirus, Chloriridovirus and Decapodiridovirus genera are larger (163–220 kbp) than those for members of the Megalocytivirus and Ranavirus genera (103–140 kbp). With the exception of SGIV, members of species within the genus Ranavirus tend to show marked sequence co-linearity and display a high level of cytosine methylation. However, even within this genus, genome inversions and deletions have been noted. Putative open reading frames (ORFs) are found on both strands of the viral genome, but overlap of coding regions is rare (Figure 3. Iridoviridae). Moreover, gene density is high, intergenic regions generally short, and genomes contain repetitive sequences. In contrast to eukaryotes, iridovirid genes lack introns and viral mRNAs lack poly[A] tails (İnce et al., 2017). However, like eukaryotes, biochemical and in silico evidence suggest the existence of viral microRNAs (miRNA) that modulate viral gene expression.
Iridovirids employ a novel replication strategy involving both nuclear and cytoplasmic stages that has been elucidated primarily through the study of FV3, the type species of the genus Ranavirus (Chinchar et al., 2009, Jancovich et al., 2015, Williams et al., 2005, Chinchar et al., 2017). Although the precise mechanism of virion entry remains to be elucidated, multiple routes have been implicated including receptor-mediated endocytosis by enveloped particles, uncoating of naked virions at the plasma membrane, pinocytosis and caveola-dependent endocytosis (Wang et al., 2014). Following uncoating, viral DNA cores enter the nucleus where first stage DNA synthesis and the synthesis of immediate early (IE) and delayed early (DE) viral transcripts take place (Figure 4. Iridoviridae). In a poorly understood process, one or more virion-associated proteins acts as a transcriptional transactivator and re-directs host RNA polymerase II to synthesize IE viral mRNAs using the methylated viral genome as template. Gene products encoded by IE (and perhaps DE) viral transcripts include both regulatory and catalytic proteins. One of these gene products, the viral DNA polymerase, catalyzes the first stage of viral DNA synthesis in which the parental viral genome serves as the template, and progeny DNA is synthesized that is genome-length to at most twice genome length. Newly-synthesized viral DNA may serve as the template for additional rounds of DNA replication or early transcription, or viral DNA may be transported to the cytoplasm where the second stage of viral DNA synthesis occurs. Second stage DNA synthesis results in the formation of large, branched concatemers. Viral DNA methylation also occurs in the cytoplasm and, although its precise role is uncertain, it is thought to protect viral DNA from virus-mediated endonucleolytic attack. Synthesis of late (L) viral transcripts takes place in the cytoplasm and is catalyzed by virus-encoded homologs of RNA polymerase II (Sample et al., 2007). Similar to other large DNA viruses, full L gene expression requires prior DNA synthesis. Thus temperature-sensitive mutants that fail to synthesize viral DNA at the non-permissive temperature, and infected cells incubated in the presence of inhibitors of DNA synthesis, display markedly reduced levels of late viral transcripts and proteins. Virion formation takes place in the cytoplasm within morphologically distinct regions termed virus assembly sites (Figure 5. Iridoviridae). Assembly sites form in cells treated with an antisense morpholino oligonucleotide (asMO) targeted against vPOL-IIα indicating that assembly site formation does not require the synthesis of late viral gene products (Sample et al., 2007). Concatemeric viral DNA is thought to be packaged into virions via a “headful” mechanism that results in the generation of circularly-permuted and terminally-redundant genomes. Following assembly, virions accumulate in the cytoplasm within paracrystalline arrays or acquire an envelope by budding from the plasma membrane (Liu et al., 2016). In the case of most vertebrate iridoviruses, the majority of virions remain cell-associated (Figure 5. Iridoviridae).
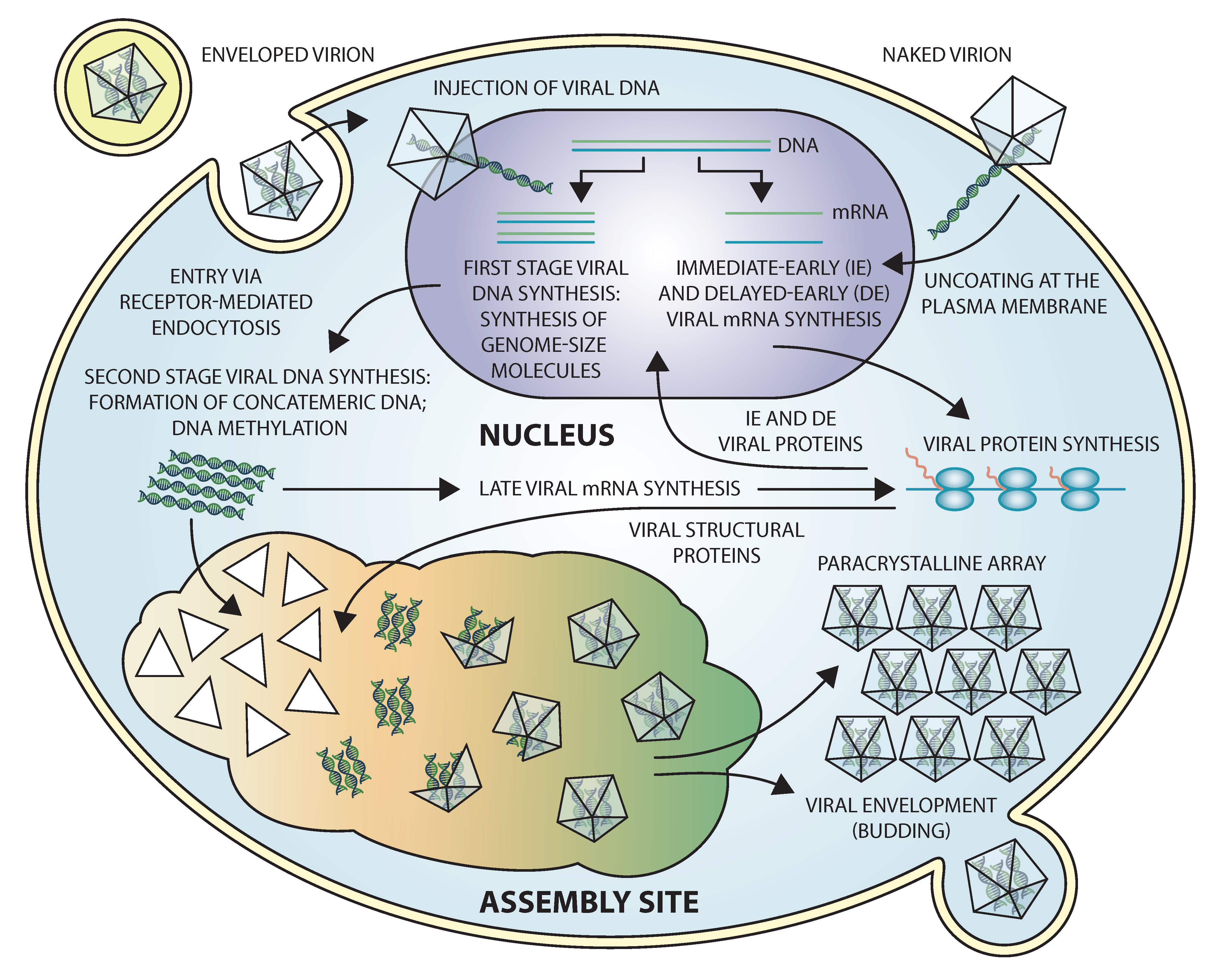 |
| Figure 4. .Iridoviridae. Replication strategy of frog virus 3 (FV3). Replication begins in the nucleus with the synthesis of early viral transcripts and the synthesis of unit size, to twice unit size, DNA genomes that are subsequently transported to the cytoplasm where a second round of DNA replication results in the formation of large concatemers. Late messages are synthesized in the cytoplasm using a multi-subunit, virus-encoded transcriptase. Virion formation takes place in morphologically distinct assembly sites and viral particles are seen within assembly sites, paracrystalline arrays, and budding from the plasma membrane. |
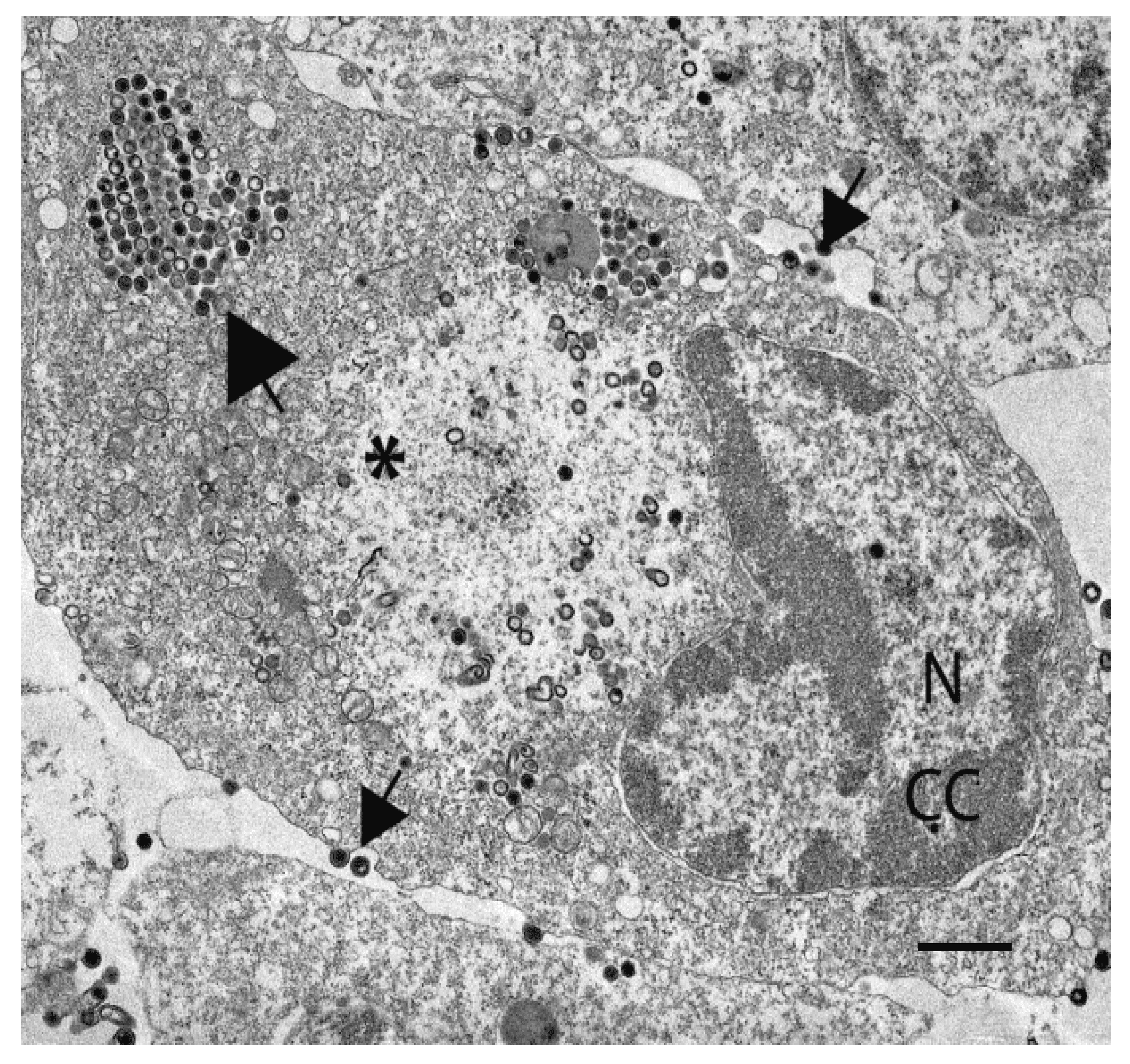 |
| Figure 5. Iridoviridae. Transmission electron micrograph of a fathead minnow cell infected with FV3. Nucleus, N, showing signs of chromatin condensation (CC); a large electron-lucent assembly site (*) containing numerous immature, empty virions, few mature virions, and devoid of cellular organelles; a paracrystalline array of virus particles (large arrow); budding virions (small arrow); scale bar = 1 µm (figure provided by VG Chinchar and RC Sample). |
Biology
Iridovirids have been isolated only from ectothermic vertebrates and invertebrates, usually associated with damp or aquatic environments, including marine and freshwater habitats. Iridovirid species vary widely in their members’ natural host range and virulence.
Invertebrate iridoviruses may be transmitted perorally, e.g. by cannibalism, as well as by endoparasitic wasps or parasitic nematodes. Infections with invertebrate iridoviruses fall into two categories: patent and covert. In the former, virus replication is extensive and infected animals display blue or green iridescence due to the presence of large numbers of virions within infected tissues. In the latter case, which is more common, lower levels of virus are present and iridescence is absent. However, despite the lower viral burden, covertly infected animals often experience reduced fitness (Williams et al., 2005).
Members of the genera Megalocytivirus and Lymphocystivirus infect only fish, whereas members of the genus Ranavirus infect fish, amphibians, and reptiles. Viruses may be transmitted experimentally by injection or bath immersion, and naturally by co-habitation and feeding, including cannibalism or scavenging of infected carcasses. Both megalocytiviruses and ranaviruses replicate within internal organs and, in the case of FV3, the kidney is a major target. Although morbidity and mortality vary depending upon the specific virus, host, and environment, mortality levels can approach 100%. In contrast, fish infected with lymphocystiviruses display wart-like growths on external surfaces (and sometimes internal organs) that are the result of greatly enlarged individual, virus-infected cells. Unlike infections with megalocytiviruses or ranaviruses that can produce mortality rates of nearly 100%, infections with lymphocystiviruses resolve spontaneously and mortality rates are generally low.
Antigenicity
Members of different genera within the family are serologically distinct from one another. However, cross-reactivity among members of different species within a given genus may occur due to sequence conservation of the MCP and other viral proteins (Ariel et al., 2010). For example, three piscine ranaviruses, EHNV, European catfish virus (ECV), and European sheatfish virus (ESV), display serological cross-reactivity with each other and with FV3 (Hedrick et al., 1992).
Studies of FV3-infected Xenopus laevis indicate a marked antibody response following secondary infection and the generation of protective neutralizing antibodies (Grayfer et al., 2015). Furthermore, consistent with a protective antibody response, inactivated virus vaccines protect against disease mediated by RSIV. In addition, in the FV3/Xenopus laevis model, T cells have been implicated in protection against viral disease (Grayfer et al., 2015, Morales and Robert 2007). However, the precise FV3 proteins that trigger a T cell response are not known.
Derivation of names
Chloro: from the Greek chloros, meaning green
Cysti: from Greek kystis, indicating a bladder or sac
Irido: from Greek iris, iridos, the goddess whose sign was the rainbow, hence iridescent, from the appearance of patently-infected invertebrates and centrifuged pellets of virions
Lympho: from Latin lympha, meaning water
Megalocyti: from the Greek, meaning “enlarged cell”
Rana: from Latin rana, “frog,” the source of the first isolates
Decapod: in reference to invertebrates such as crayfish, crabs, prawns, and shrimp (order Decapoda, class Malacostraca, subphylum Crustacea) with five pairs of walking legs.
Alpha/Beta: Greek letters indicating the chronological order in which iridovirid pathogens of vertebrates and insects were identified
Subfamily demarcation criteria
The two subfamilies within the family Iridoviridae are distinguished by their primary hosts. Members of the Alphairidovirinae infect mainly ectothermic vertebrates (bony fish, amphibians, and reptiles), whereas members of the Betairidovirinae infect mainly invertebrates (insects and crustaceans). Furthermore members of the latter subfamily tend to have larger genomes with a lower G+C content, and lack extensive DNA methylation.
Genus demarcation criteria
Members of the six genera within the family are distinguished by the level of nucleotide/amino acid sequence identity, host range, G+C content, phylogenetic relatedness, genome co-linearity, disease manifestations and antigenicity. Members of species within the same genus generally show greater than 50% sequence identity within a common set of 26 core genes.
Relationships within the family
Phylogenetic analyses (Figure 6. Iridoviridae), based on individual genes, or a concatenated set of 26 genes common to all members of the family, support the division of the family into two subfamilies and six genera (Chinchar et al., 2011, Jancovich et al., 2015). Viruses within a genus generally share greater than 50% identity at the nucleotide and amino acid levels. Moreover, viruses displaying greater than 95% sequence identity are considered to be isolates/strains of the same viral species. For example, the ranaviruses FV3, Bohle iridovirus (BIV), tiger frog virus (TFV), and Rana gyrilo virus (RGV) show >95% sequence identity and a similar genomic organization and are viewed as isolates of the same viral species, Frog virus 3. In contrast, FV3, ATV, and EHNV show >90% sequence identity, but differ in host range and genomic organization and thus are classified as members of separate species. Furthermore, members of the six currently recognized species within the genus Ranavirus fall into four distinct phylogenetic clusters (Figure 6. Iridoviridae), while megalocytiviruses fall into two clusters and lymphocystiviruses fall into three clusters.
As additional viruses are isolated and genomes sequenced, the status of viral species within these genera will be re-evaluated, taking into account additional criteria such as host range, gene order, the presence or absence of specific genes, nucleotide and amino acid sequence identity, and antigenicity.
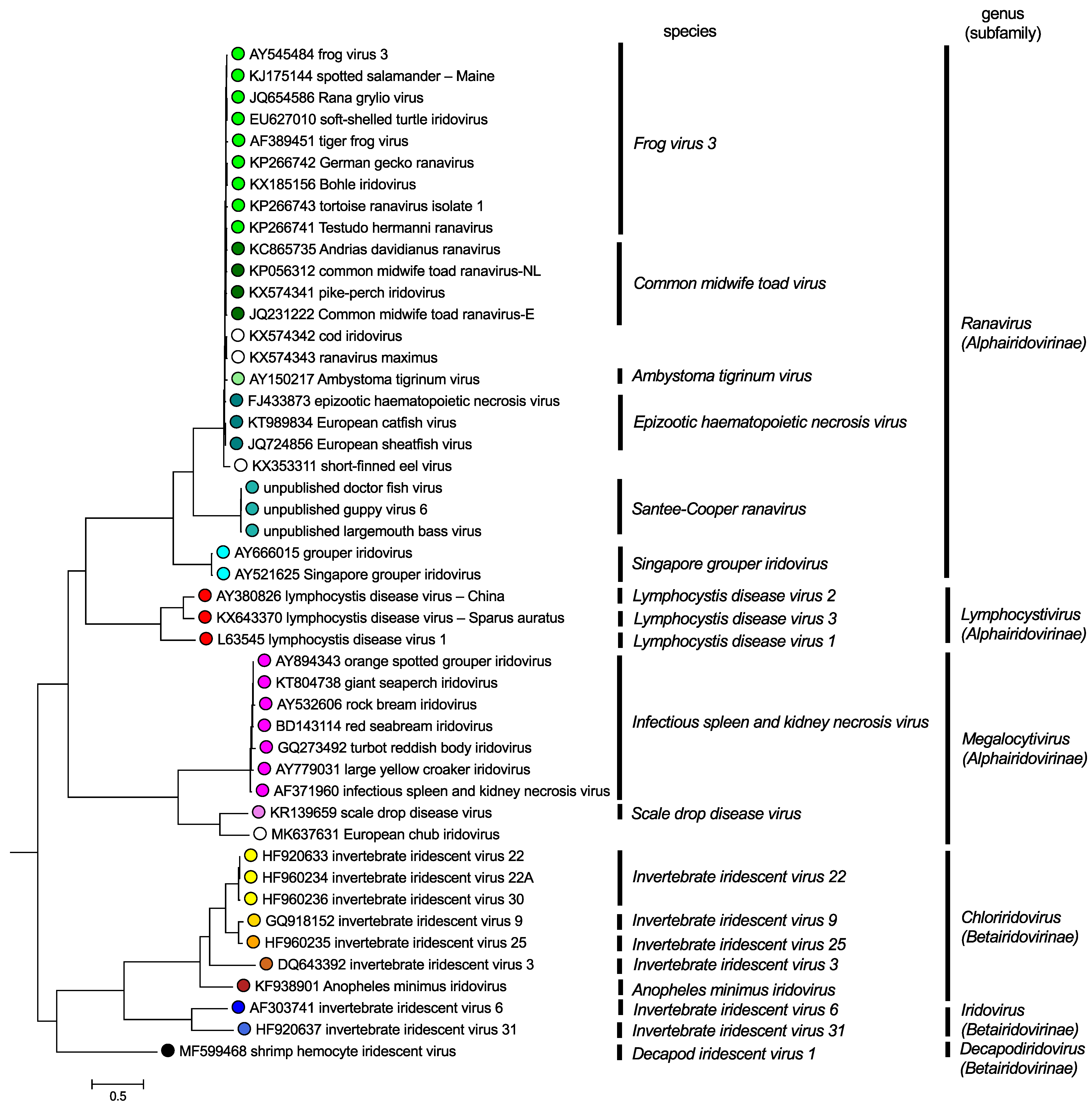 |
Figure 6.Iridoviridae: Phylogenetic analysis of family Iridoviridae. The tree was constructed using maximum likelihood analysis in IQTREE and the concatenated amino acid (aa) sequences of 24 core genes (20,023 aa characters including gaps) from 47 completely sequenced iridovirus genomes. The tree was midpoint rooted and branch lengths are based on the number of inferred substitutions, as indicated by the scale bar. All nodes were supported by bootstrap values greater than 70% with the vast majority displaying values of > 99%. (analysis performed by K Subramaniam and TB Waltzek). The isolates cod iridovirus, ranavirus maximus, short-finned eel virus are unclassified viruses in the genus Ranavirus, and European chub iridovirus is an unclassified virus in the genus Megalocytivirus. This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. |
Relationships with other taxa
Iridovirid homologs of D5 ATPase, A32 ATPase, the A1L/VLTF2 transcription factor, MCP and viral DNA polymerase display amino acid sequence similarity with African swine fever virus and members of the families Ascoviridae, Phycodnaviridae, Mimiviridae and Poxviridae (Boyer et al., 2009). These relationships have prompted the suggestion that these viruses share a common origin and constitute members of a (Colson et al., 2013) higher level taxon. Likewise, using a concatenated set of 9 genes common to iridovirids, ascoviruses and marseilleviruses, the close relationship between ascoviruses and invertebrate iridescent viruses has been confirmed, suggesting that ascoviruses emerged recently and share a common ancestor with IIV6 and IIV3 (Piegu et al., 2015). However, given the markedly different morphologies and replication strategies of ascoviruses and iridoviruses (Federici et al., 2009), additional analysis will be required to resolve the complicated phylogeny of members of these two distinct, yet related, families.
Related, unclassified viruses
| Virus name | Accession number | Virus abbreviation | Genome (bp) | ORFs | %GC |
| Erythrocytic necrosis virus | KJ756346 | ENV | ND1 | ND | ND |
| Daphnia iridescent virus 1 | LS484712 | DIV1 | 288,000 | 367 | 39 |
Virus names and abbreviations are not official ICTV designations.
1 The complete genomic sequence of ENV has not yet been determined. Listed is the partial sequence of the viral DNA-dependent RNA polymerase of the herring isolate. Preliminary evidence suggests that ENV and DIV1 might be members of new genera within the family.