

Family: Dicistroviridae
Steven M Valles, Yanping Chen, Andrew E Firth, Diego M. A Guérin, Yoshifumi Hashimoto, Salvador Herrero, Joachim R de Miranda and Eugene Ryabov
The citation for this ICTV Report chapter is the summary published as Valles et al., (2017):
ICTV Virus Taxonomy Profile: Dicistroviridae, Journal of General Virology, 98: 355–356.
Corresponding author: Steven M. Valles ([email protected])
Edited by: Nick J. Knowles and Peter Simmonds
Posted: March 2017, updated May 2025
PDF: ICTV_Dicistroviridae.pdf (2017 version)
Summary
Dicistroviridae is a family of small non-enveloped viruses with RNA genomes of approximately 8–10 kilobases (Table 1 Dicistroviridae). All members infect arthropod hosts with some having devastating economic consequences, such as acute bee paralysis virus, Kashmir bee virus, and Israeli acute paralysis virus in domesticated honeybees, and taura syndrome virus and mud crab virus in the seafood industry. Conversely, host specificity and other desirable traits make several members of this group amenable to development as biopesticides for insect control, such as Solenopsis invicta virus 1 against fire ants, and Triatoma virus against triatomine bugs that vector Chagas disease.
Table 1 Dicistroviridae. Characteristics of members of the family Dicistroviridae
| Characteristic | Description |
| Typical member | cricket paralysis virus (AF218039), species Cripavirus grylli |
| Virion | Non-enveloped, 30 nm diameter virions |
| Genome | 8–10 kb of positive-sense, non-segmented RNA |
| Replication | Cytoplasmic within viral replication complexes formed from a variety of host cellular membranes |
| Translation | Directly from genomic RNA, initiated at internal ribosome entry site elements in the 5′-NCR and intergenic untranslated region |
| Host Range | Arthropoda |
| Taxonomy | Realm Riboviria, kingdom Orthornavirae, phylum Pisuviricota, class Pisoniviricetes, order Picornavirales; includes the genera Aparavirus, Cripavirus, and Triatovirus each containing several species for a total of 16 species |
Virion
Morphology
Virions are roughly spherical with a particle diameter of approximately 30 nm and no envelope (Figure 1 Dicistroviridae). The virions exhibit icosahedral, pseudo T=3 symmetry and are composed of 60 protomers, each comprising of a single molecule of VP1, VP2 and VP3 (Figure 1 Dicistroviridae). A smaller protein, VP4, is also present inside the virion and in contact with the genome. In cricket paralysis virus and Israeli acute paralysis virus, VP4 is structured on the internal surface of the 5-fold axis below VP1, but in contrast is disordered in Triatoma virus (Sanchez-Eugenia et al., 2015, Mullapudi et al., 2016).
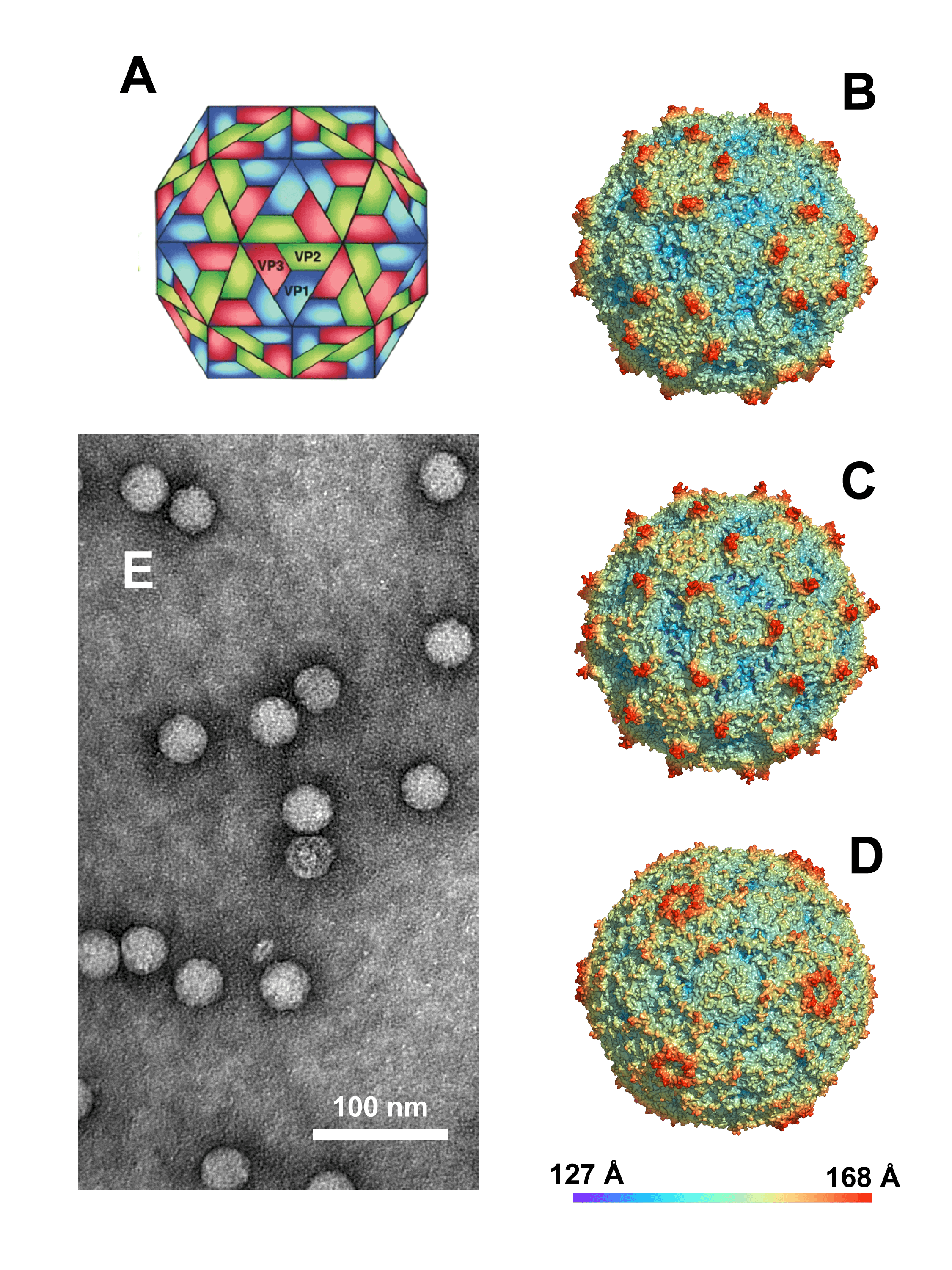 |
| Figure 1 Dicistroviridae. (A) Diagram illustrating the packing of surface proteins VP1, VP2, and VP3. The smallest structural protein, VP4, is found in the capsid interior. Renderings (courtesy of A.E. Mechaly) of (B) Triatoma virus, (C) Israeli acute bee paralysis virus, and (D) cricket paralysis virus (color scale indicates distance from the particle center). (E) Negative-contrast electron micrograph of purified Triatoma virus (courtesy of G.A. Marti). |
Physicochemical and physical properties
Virions are stable in acidic conditions (to pH 3.0). Alkaline conditions (pH > 8.5) trigger virion disassembly in Triatoma virus (Snijder et al., 2013). Virions have a buoyant density of 1.34–1.39 g ml−1 in CsCl in the pH range of 7 to 9 and sedimentation coefficients of between 153 and 167 S. However, physicochemical properties have not been fully established for all members of the family.
Nucleic acid
Virions contain a single molecule of infectious, linear, positive-sense, single-stranded RNA of approximately 8–10 kilobases with a GC content ranging from 34 to 45%. RNA constitutes 34% (median) of the virion mass. A small genome-linked virus protein (VPg), is covalently attached to the 5′-end of the genome. A 5′-non coding region (NCR) is followed by two non-overlapping open reading frames (ORF 1 and ORF 2) of approximately 5,500 and 2,700 nucleotides, respectively. The ORFs are separated by an intergenic untranslated region (IGR) of about 170–530 nucleotides that contains an internal ribosome entry site (IRES). An NCR is also found at the 3′-end of the genome. The 3′-terminus of the viral RNA genome is polyadenylated. The length of the NCRs at each end of the genome is variable. A clone of the cricket paralysis virus has been produced and found to be infectious (Kerr et al., 2015).
Proteins
Proteins account for approximately 70% of the virion mass. The approximately 200 kDa nonstructural polyprotein and 100 kDa structural polyprotein are encoded by ORF 1 and ORF 2, respectively. Virions contain three major structural (capsid) viral proteins, VP1, VP2 and VP3. The size of these capsid proteins ranges from 24 to 40 kDa; an exception is taura syndrome virus in which VP1 is 55 kDa. A fourth smaller capsid protein (VP4) of around 4.5–9 kDa also has been reported in some species. In most species a protein precursor (VP0) is initially synthesized and subsequently cleaved to yield capsid proteins VP3 and VP4.
Capsid features that differentiate the dicistroviruses from those of enteroviruses are: i) the absence of a depression (canyon) at the capsid surface; ii) the lack of the characteristic enterovirus pocket factor found in the interior of the VP1-barrel; and iii) the swapping of the VP2 N-terminus. Conservation of the amino acids comprising the strand swap suggests that this feature is present across the entire family.
Genome organization and replication
The RNA genome is monopartite and contains two main non-overlapping ORFs that are separated and flanked by NCRs (Figure 2 Dicistroviridae). The 5′-proximal and 3′-proximal ORFs encode non-structural and structural protein precursors, respectively. Components of the non-structural polyprotein include an RNA-dependent RNA polymerase (RdRP), cysteine protease, RNA helicase and one or more copies of a VPg protein. VPg is covalently linked to the 5′ end of the genome.
 |
| Figure 2 Dicistroviridae. Genome structure of cricket paralysis virus. The RNA genome contains two non-overlapping ORFs separated by an intergenic region (IGR). The 5′-proximal ORF encodes the nonstructural proteins: RNA helicase (Hel), cysteine protease (Pro), and RNA-dependent RNA polymerase (RdRP). The structural proteins are encoded by the 3′-proximal ORF and are expressed as a polyprotein that is subsequently processed to yield three major structural proteins (VP1, VP2 and VP3). A fourth structural protein, VP4, is the N-terminal extension of VP3, which is cleaved from the precursor VP0. This small peptide is considered to be involved in membrane alterations required for genome transfer to the cytoplasm of host cells during infection. Indeed, recent independent studies have shown that recombinant VP4 from both cricket paralysis virus and Triatoma virus forms pores in model membranes (Sanchez-Eugenia et al., 2015, Kerr et al., 2015). Distinct internal ribosome entry sites (IRES) are located in the 5′-NCR (labelled 5′-UTR) and IGR. The genome has a small peptide covalently linked to the 5′-end (genome-linked virus protein, VPg) and a 3′-polyadenylated terminus. |
Replication occurs exclusively in the cytoplasm of infected cells. Cap-independent translation of dicistrovirus genomes proceeds directly from two distinct IRES elements located within the 5′-NCR and the IGR. These IRES elements permit production of non-structural proteins early in the infection process before host translation mechanisms are inhibited, and excess molar quantities of structural proteins from the IGR IRES when capsid proteins are required later. Unusually, the IGR IRES directs translation initiation at a 3′-adjacent non-AUG codon and in the absence of all canonical initiation factors. The conserved 3-dimensional structure is crucial to the IGR IRES function (Jan 2006). Pseudo-knot and stem-loop structures in the IGR IRES are highly conserved across all members of the family and facilitate interactions with the ribosome (Nakashima and Uchiumi 2009). The 5′-NCR IRES is not obviously conserved in sequence or structure across the group. Translation activity of the IGR IRES is comparatively greater than that of the 5′-NCR IRES (Khong et al., 2016).
Biology
All members of the family infect arthropods (Bonning and Miller, 2010). Most members of the family are distributed widely in nature. Aggregates and crystalline arrays of virus particles have been observed in the cytoplasm of infected cells. Dicistrovirus infections vary considerably in virulence and pathogenicity, ranging from inapparent to lethal. Transmission by ingestion and spread from the alimentary canal feature prominently in dicistrovirus infection acquisition and transmission. Most of the dicistroviruses exhibit tissue tropism to parts of the alimentary canal, often replicating in epithelial cells of the gut and subsequently shedding virus particles into the gut lumen where virus accumulates in the faeces (an often important source of infectious virus). An additional (and unique) route for horizontal transmission is observed in some of the honeybee viruses (acute bee paralysis virus, Kashmir bee virus, Israeli acute bee paralysis virus and black queen cell virus). In these, infections of the adult brain result in virus shedding via glandular secretions (e.g., royal jelly), which are fed to honeybee larvae facilitating intra-colonial dispersal of the virus. </p> <p> Nervous tissue, fat body, epidermal cells, and gonads may also support replication of dicistroviruses. Vertical virus transmission (transovarial or transovum) is also exhibited (acute bee paralysis virus, Triatoma virus, Plautia stali intestine virus, Solenopsis invicta virus 1). In honeybees, acute bee paralysis virus, Kashmir bee virus, and Israeli acute paralysis virus are transmitted by parasitic mites (<em>Varroa</em> and <em>Tropilaelaps</em> genera) resulting in highly dysfunctional and short-lived adult bees and colony mortality. Host specificity is largely unexplored. Solenopsis invicta virus 1 appears limited to the <em>Solenopsis</em> genus of ants. Conversely, the honeybee viruses, black queen cell virus, acute bee paralysis virus, Kashmir bee virus, and Israeli acute bee paralysis virus also infect <em>Bombus</em> bee species (<a href=" https:="" www.ncbi.nlm.nih.gov="" pubmed="">McMahon et al., 2015), affecting colony development and reproductive success (Meeus et al., 2014).
Antigenicity
Many members of the family are serologically distinguishable. Studies have shown that acute bee paralysis virus, Kashmir bee virus, and Israeli acute paralysis virus are serologically related (Allen and Ball 1995).
Derivation of names
Aparavirus: from acute bee paralysis virus, a virus in the genus
Cripavirus: from cricket paralysis virus, a virus in the genus
Dicistroviridae: derived from the characteristic dicistronic arrangement of the genome
Triatovirus: from Triatoma virus, a virus in the genus
Genus demarcation criteria
Formal genus demarcation criteria have not yet been established. However, different genera display unique sequence and topological characteristics in the IGR IRES and form separate groups on phylogenetic analysis (Figure 3A Dicistroviridae).
Relationships within the family
Phylogenetic analysis of structural polyprotein amino acid sequences indicates that cripaviruses, aparaviruses, and triatoviruses form distinct groups in the family (Figure 3A. Dicistroviridae). Genus Aparavirus, however, separates further into two distinct divergent clades comprising, respectively, the insect-infecting members and the crustacean-infecting members. Differences in host, details of the IGR-IRES, and presence of an additional ORF overlapping the 5′-region of the structural ORF in the former that is not conserved in the latter indicate that genus Aparavirus is likely to require revision to form two genera. Phylogenetic analysis of nonstructural polyprotein amino acid sequences also supports four distinct clades (cripaviruses, triatoviruses, and two distinct groups of the aparaviruses) (Figure 3B Dicistroviridae).
 |
| Figure 3 Dicistroviridae. Mid-point rooted phylogenetic tree constructed from full-length polyprotein amino acid sequences of formally accepted dicistrovirus species and related unclassified virus sequences using MrBayes. While we include metagenome-derived sequences (black text) to provide a more comprehensive phylogeny of the group, we caution that the names given to such sequences are tentative and may not accurately reflect the true host or biology. The currently defined genera are annotated at right and members of the current species are indicated by blue font. (A) Structural polyprotein tree. (B) Nonstructural polyprotein tree. Node labels indicate posterior probabilities. These phylogenetic trees and corresponding sequence alignments are available to download from the Resources page. |
Relationships with other taxa
Members of the family Dicistroviridae are similar to viruses in the Iflaviridae, Picornaviridae, Marnaviridae and Secoviridae families, which together form the order Picornavirales. The genomes of viruses of these taxa are positive-sense ssRNAs with a 5′-linked VPg and a poly (A) tail, and are translated into autoproteolytically processed polyprotein(s). Nonstructural proteins contain sequence motifs for helicase (Hel), 3C-like cysteine proteinase (Pro) and RNA-dependent RNA polymerase (RdRP) with the characteristic gene order: (Hel)-(Pro)-(RdRP). Virions contain capsid proteins organized in a module containing three related jelly-roll domains which form non-enveloped, isometric particles of pseudo T=3 symmetry and ~30 nm diameter.