

Family: Aspiviridae
María Laura García, Elena Dal Bó, John V. da Graça, Selma Gago-Zachert, John Hammond, Pedro Moreno, Tomohide Natsuaki, Vicente Pallás, Jose A. Navarro, Carina A. Reyes, Gabriel Robles Luna, Takahide Sasaya, Ioannis E. Tzanetakis, Anna María Vaira and Martin Verbeek
The citation for this ICTV Report chapter is the summary published as García et al., (2017):
ICTV Virus Taxonomy Profile: Ophioviridae, Journal of General Virology, 98: 1161–1162.
Corresponding author: María Laura García ([email protected])
Edited by: Hélène Sanfaçon and Michael J. Adams
Posted: June 2017 (as Ophioviridae), updated July 2019
PDF: ICTV_Aspiviridae.pdf
Summary
The Aspiviridae (formerly Ophioviridae) is a family of filamentous plant viruses, with single-stranded negative-sense, and possibly ambi-sense, RNA genomes of 11.3–12.5 kb divided into 3–4 segments (Table 1. Aspiviridae). Virions are naked filamentous nucleocapsids, non-enveloped, in the shape of kinked circles of at least two different contour lengths. Since only one genus is currently recognized, the family description corresponds to that of the genus Ophiovirus; the type species is Citrus psorosis ophiovirus. The family includes seven species, of which four are known to be soil-transmitted. Natural hosts of aspiviruses include trees, shrubs, vegetables and bulbous or corm-forming ornamentals, both monocots and dicots.
Table 1. Aspiviridae. Characteristics of members of the family Aspiviridae.
| Characteristic | Description |
| Typical member | citrus psorosis virus P-121 (RNA1 AY654892; RNA2: AY654893; RNA3: AY654894), species Ophiovirus citri |
| Virion | Non-enveloped, naked filamentous nucleocapsids about 3 nm in diameter, forming kinked circles of at least two different contour lengths (700 nm and about 2000 nm). Pseudo-linear duplex structures are about 9–10 nm in diameter |
| Genome | 11.3–12.5 kb of negative-sense, segmented RNA (3 or 4 segments) |
| Replication | Not characterized |
| Translation | From mRNAs, which are complementary to the vRNAs |
| Host Range | Citrus, blueberry, pittosporum, lettuce, sowthistle, tulip, ranunculus, anemone, lachenalia and freesia |
| Taxonomy | Realm Riboviria, kingdom Orthornavirae, phylum Negarnaviricota, subphylum Haploviricotina, class Milneviricetes and order Naedrevirales; one genus, Ophiovirus, including 8 species |
Virion
Morphology
The particles are naked filamentous nucleocapsids about 3 nm in diameter (Figure 1. Aspiviridae), forming kinked (probably internally coiled) circles of at least two different contour lengths, the shortest being about 700 nm. The circles (open form) can collapse to form pseudo-linear duplex structures of 9–10 nm in diameter (collapsed form) (Milne et al., 1996).
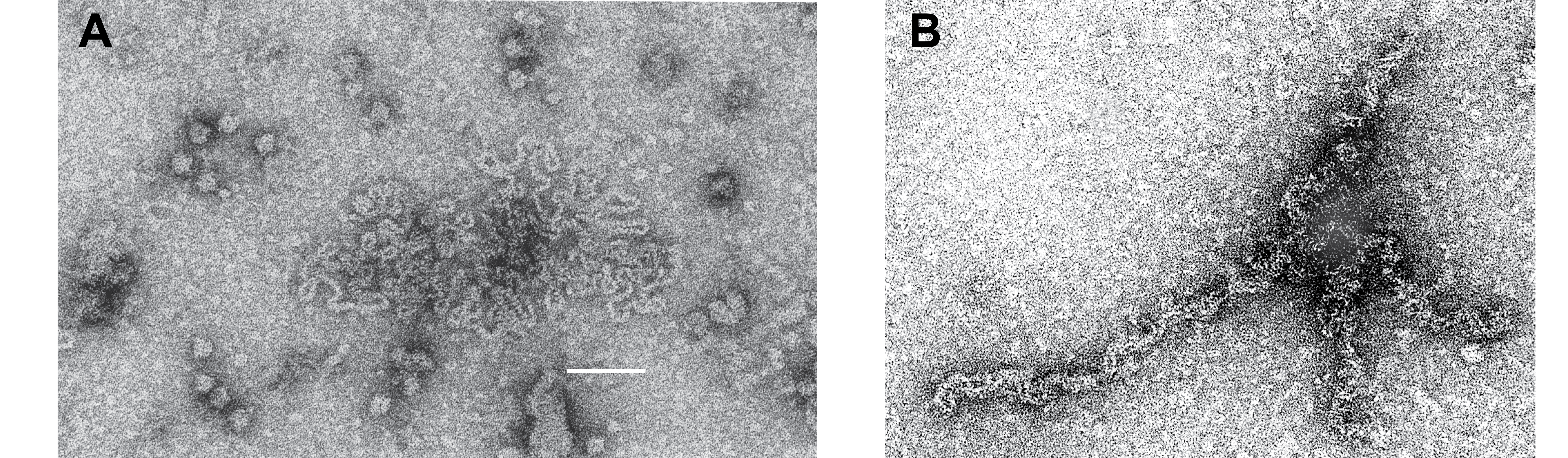 |
| Figure 1. Aspiviridae. Negative-contrast electron micrograph (uranyl acetate) of aspivirus particles. The bar represents 100 nm. (A) Open form of the large particle (citrus psorosis virusV), and (B) collapsed form (freesia sneak virus) (Courtesy of R. G. Milne). |
Physicochemical and physical properties
The relative molecular masses and sedimentation coefficients of virions are unknown. The particles are unstable in CsCl but the buoyant density in cesium sulfate has been determined to be 1.22 g cm−3 for both ranunculus white mottle virus [RWMV, Rn3 isolate], and Mirafiori lettuce big-vein virus [MLBVV, I-47 isolate] (Roggero et al., 2000, Vaira et al., 1997). The particles are unstable between pH 6 and 8. Viruses lose infectivity in crude sap held at 50 °C for 10 min (40–45 °C for tulip mild mottle mosaic virus, TMMMV) (Morikawa et al., 1995). Virus structures are resistant to limited treatment with organic solvents and nonionic or zwitterionic detergents.
Nucleic acid
Aspiviruses have a single-stranded negative-sense, and possibly ambi-sense RNA genome (11.3–12.5 kb in total) divided into 3 or 4 segments. The virions encapsidate both the viral and viral complementary RNAs (vRNA and vcRNA), but a larger amount of vRNA (negative-sense) is detected in purified virion preparations (Roggero et al., 2000, Garcia et al., 1991, Naum-Ongania et al., 2003, Sánchez de la Torre et al., 1998, van der Wilk et al., 2002). As virions appear circularized, the presence of panhandle structures has been suggested; however, significant sequence complementarity was not found between the 5′- and the 3′-terminal sequences of genomic RNAs of citrus psorosis virus (CPsV), MLBVV or blueberry mosaic associated virus (BlMaV) (Naum-Ongania et al., 2003, van der Wilk et al., 2002, Thekke-Veetil et al., 2014). At the 5′- and 3′-ends of MLBVV genomic RNAs, there are orthomyxovirus-like palindromic sequences that could fold into a symmetrically hooked conformation. These structures could not be predicted for other sequenced members.
RNA1 is 7.5–8.2 kb, RNA2 is 1.6–1.9 kb and RNA3 is 1.4–1.5 kb. A fourth genomic RNA of about 1.4 kb has been reported for MLBVV and lettuce ring necrosis virus (LRNV). In the case of CPsV, RWMV, freesia sneak virus (FreSV) and BlMaV RNA4 was not found; no information is available for TMMMV (Vaira et al., 1997, Morikawa et al., 1995, Naum-Ongania et al., 2003, Sánchez de la Torre et al., 1998, Thekke-Veetil et al., 2014, Sánchez de la Torre et al., 2002, Torok and Vetten 2002, Torok and Vetten 2003, Vaira et al., 2003, Vaira et al., 2009, Vaira et al., 1996).
Proteins
Virions consist of a single type of coat protein (CP) subunit of 43–50 kDa, varying in mass according to species and isolate (Roggero et al., 2000, Vaira et al., 1997, Morikawa et al., 1995, Sánchez de la Torre et al., 1998, van der Wilk et al., 2002, Torok and Vetten 2003, Derrick et al., 1988, Garcia et al., 1994, Jeong et al., 2014, Navas‐Castillo et al., 1993).
Genome organization and replication
The genome of aspiviruses consists of three or four individually encapsidated vRNA segments (Figure 2. Aspiviridae) although vcRNAs are also encapsidated (Vaira et al., 1997, Naum-Ongania et al., 2003, Sánchez de la Torre et al., 1998, van der Wilk et al., 2002, Thekke-Veetil et al., 2014, Sánchez de la Torre et al., 2002, Vaira et al., 2003, Martin et al., 2005). The vcRNA1, the longest RNA segment (7.5–8.2 kb), contains two ORFs. The ORF located at its 5′-end encodes a protein with a predicted molecular mass of 22–25 kDa. The 24K ortholog of CPsV localizes at the nucleus and was reported to be involved in miRNA misprocessing in citrus (Reyes et al., 2016), and to possess RNA silencing suppressor activity (Robles Luna et al., 2017). Separated by an intergenic region of about a hundred nucleotides, a second ORF in the same strand encodes a protein (261–280 kDa) that contains the core polymerase module with the characteristic five conserved motifs (A–E) of the RNA-dependent RNA polymerases (RdRP). The SDD sequence, a signature for segmented negative-stranded RNA viruses (Orthomyxoviridae, Arenaviridae and Bunyaviridae) occurs in motif C of the RdRP (Naum-Ongania et al., 2003). Since no other segmented RNA viruses have two ORFs coded in the vcRNA of the larger segment (RNA1), this genomic organization is a particular feature of members of the Aspiviridae family.
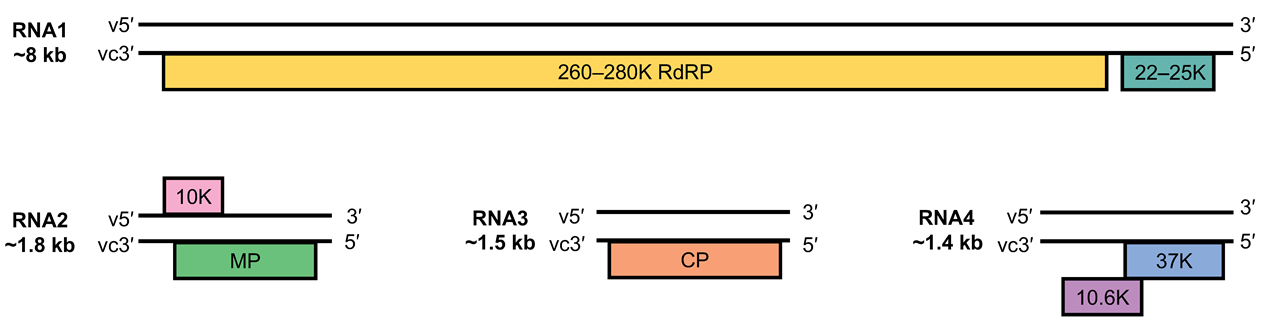 |
| Figure 2. Aspiviridae. Genome organization of members of the genus Ophiovirus. Using the example of Mirafiori lettuce big-vein virus (modified from (van der Wilk et al., 2002)), boxes represent ORFs and the length of the RNA segments and the predicted sizes of the ORF products are indicated. v, viral RNA; vc, viral complementary RNA. RNA4 is not reported for all ophioviruses. RdRP, RNA polymerase; MP, movement protein; CP, coat protein. |
The vcRNA2 (1.6–1.9 kb) of CPsV, LRNV, FreSV and BlMaV contains a single ORF, whereas for MLBVV a second ORF in vRNA2 encoding a putative protein of 37–38 kDa of unknown function has been reported. This would make MLBVV the sole ophiovirus with ambisense RNA2 (van der Wilk et al., 2002). The vcRNA2 encodes the cell-to-cell movement protein (MP) of 50-58 kDa, which has been characterized for CPsV (54K) and MLBVV (55K) (Hiraguri et al., 2013, Robles Luna et al., 2013). The MP is also involved in RNA silencing suppression activity (Robles Luna et al., 2017). The vcRNA3 codes for the CP (Roggero et al., 2000, Sánchez de la Torre et al., 1998, Vaira et al., 2011); the CP molecules of MLBVV and CPsV can interact among themselves, and localize in the cytoplasm of N. benthamiana epithelial cells, when expressed ectopically (Peña et al., 2012). RWMV CP accumulates in the cytoplasm of parenchyma cells (Roggero et al., 2000). The CPs of MLBVV and CPsV interact with the virus-specific MP in the cytoplasm suggesting a potential role of the CP in ophiovirus movement (Robles Luna et al., 2013).
RNA4 (ca. 1.4 kb) has been identified only from MLBVV and LRNV (Roggero et al., 2000, Torok and Vetten 2003). Whereas the vcRNA4 of LRNV potentially encodes only one protein (ca. 38 kDa) of unknown function, that of MLBVV contains two ORFs overlapping by 38 nt, potentially encoding a 37 and 10.6 kDa proteins. The second ORF lacks an initiation codon and is proposed to be expressed by a +1 translational frameshift (van der Wilk et al., 2002).
Nuclear localization signals (NLS) have been reported for the CPsV, MLBVV, BlMaV and RWMV polymerases, for the MPs of CPsV and MLBVV, and for the 23K protein of BlMaV (Naum-Ongania et al., 2003, van der Wilk et al., 2002, Thekke-Veetil et al., 2014, Sánchez de la Torre et al., 2002).
For CPsV and BlMaV, the 5′-ends of the vcRNA1, vcRNA2 and vcRNA3 are identical, GATAC(T)7, but the 3′-ends are all different. For MLBVV, the respective 5′- and 3′-ends are conserved among the four viral RNA segments. In addition, the vRNA terminal ends are able to anneal and fold into structures resembling the ‘corkscrew’ conformation of the RNA termini of the Orthomyxoviridae (van der Wilk et al., 2002).
For BlMaV, there are indications of natural reassortments between RNA2 and RNA3 (Thekke-Veetil et al., 2015). Natural reassortments between RNA1 and RNA3 of CPsV have been suggested (Martin et al., 2006), and genome compatibility, between the bottom (RNA1) and top (RNA2 and RNA3) components of two CPsV isolates, has been observed in the experimental host Chenopodium quinoa (Garcia et al., 1993).
Biology
Ophioviruses can be mechanically transmitted to a limited range of test plants, the most used being Nicotiana benthamiana and N. occidentalis P1, inducing local lesions and systemic mottle, and Chenopodium quinoa producing local lesions. The natural hosts of CPsV, RWMV, MLBVV and LRNV are dicotyledonous plants of widely differing taxonomy. In contrast, TMMMV and FreSV infect monocots. CPsV is commonly transmitted by vegetative propagation of the host. Although natural dispersion of psorosis disease has been observed in Texas and Argentina (Danós 1990, Timmer and Garnsey 1980) no natural vector has been identified, since limited vector-transmission experiments have been performed (Beñatena and Portillo 1984, Palle et al., 2004). No vector is known for either RWMV or BlMaV. The zoospores of Olpidium spp. transmit MLBVV, TMMMV, LRNV and FreSV (Lot et al., 2002, Meekes and Verbeek 2011, Navarro et al., 2004, Sasaya and Koganezawa 2006, Vaira et al., 2006). MLBVV has been shown to be present in the weed Sonchus oleraceus which can act as a natural reservoir for lettuce infection by means of the fungus vector (Navarro et al., 2005).
CPsV has a wide geographical distribution in Citrus spp. and citrus relatives in the Americas, in the Mediterranean basin, New Zealand and South Africa (Quemin et al., 2011, Roistacher 1993, Sippel et al., 2015).
RWMV has been reported in two species of the family Ranunculaceae from Northern Italy (ranunculus and anemone hybrids) (Vaira et al., 1997, Vaira et al., 2003), in lettuce in France and Germany (Torok and Vetten 2010) and recently in the evergreen shrub Pittosporum tobira in Southern Italy (Morelli et al., 2015).
FreSV has been reported in Europe, South Africa, North America, South Korea and New Zealand (Vaira et al., 2009, Jeong et al., 2014, Vaira et al., 2006, Pearson et al., 2009, Vaira et al., 2007, Verbeek et al., 2004) in freesia (Freesia refracta hyb., Iridaceae) and in lachenalia (Lachenalia hyb., Hyacinthaceae).
TMMMV has been reported in tulips (Tulipa hyb., Liliaceae) in Japan (Morikawa et al., 1995). MLBVV is the causal agent of big-vein symptoms (zones cleared of chlorophyll, parallel to the veins) in lettuce; this widespread and damaging disease probably occurs worldwide.
LRNV is closely associated with lettuce ring necrosis disease in The Netherlands and Belgium (Torok and Vetten 2002). BlMaV has been reported in blueberry (Vaccinium spp., Ericaceae) in several regions of North America as well as several other parts of the globe including South America, Europe, New Zealand, South Africa and Japan (Isogai et al., 2016, Martin et al., 2012).
Antigenicity
The CP is the only significant antigenic element. In Western blots, the CPs of RWMV, TMMMV, MLBVV and LRNV, but not CPsV, appear to be serologically related to varying degrees (Roggero et al., 2000).
Derivation of names
Aspiviridae: from Latin aspis, “viper, snake”, referring to the morphology of virions.
Ophiovirus: from the Greek ophis, “snake”, referring to the snaky appearance of the virion (Garcia et al., 1994).
Relationships within the family
Ophiovirus-specific primers, based on a highly conserved sequence of RNA1, have been tested in RT-PCR with all ophiovirus species. In all cases, a 136 bp fragment was amplified (Vaira et al., 2003). Phylogenetic analysis of the coat protein from one isolate for each species (Figure 3. Aspiviridae) supported the positions of each species and indicated a closer relationship between MLBVV and TMMMV, as already suggested by coat protein serological tests. BlMaV and CPsV cluster in a distinct clade and seem more distantly related to the other ophioviruses by molecular characteristics. These differences, if reinforced, might also lead to the re-assignment of the existing species into two separate genera in the family. Analysis of the conserved RdRP motifs show again a distinct clade of CPsV and BlMaV, as well as higher identities between the orthologous CP and MP, with respect to the rest of the ophioviruses (Thekke-Veetil et al., 2015).
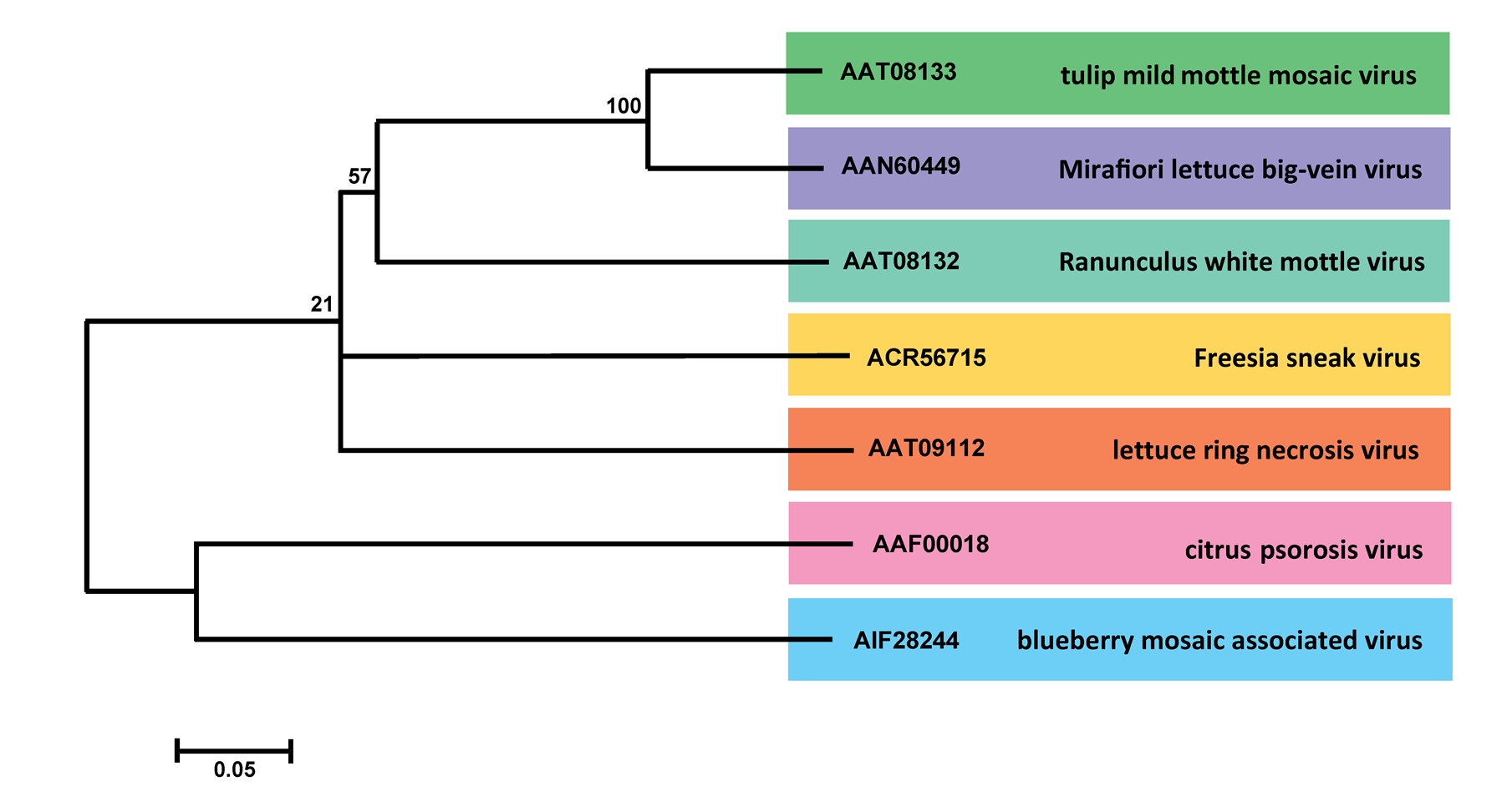 |
| Figure 3. Aspiviridae. Unrooted phylogenetic tree based on the predicted coat protein amino acid sequences encoded by RNA3 of isolates of 7 ophiovirus species. Alignment was obtained using ClustalW, and analyzed by the neighbor-joining method, with 1000 bootstrap replicates. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test is shown next to the branches (when >50%). The evolutionary distances were computed using the p-distance method. This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. |
Relationships with other taxa
The higher taxonomy of the Aspiviridae family is: realm Riboviria, phylum Negarnaviricota, subphylum Haploviricotina, class Milneviricetes and order Serpentovirales (Figure 4). Ophiovirus virion morphology resembles that of polyploviricotene viruses in the genus Tenuivirus, family Phenuiviridae, and the internal nucleocapsid component resembles that of members of the family Tospoviridae. Unlike tenuiviruses, ophioviruses do not infect plants in the Gramineae, and unlike members of the family Tospoviridae, there is no evidence of enveloped virions. Moreover, ophioviruses do not have the conserved identical nucleotides at the genomic RNA termini that are typical of these two families. MLBVV appears to have a terminal “corkscrew”-like conformation similar to that in viruses in the Orthomyxoviridae. RdRP aa sequences show ophioviruses to be similar to members of the haploviricotine families Paramyxoviridae, Pneumoviridae, Rhabdoviridae, Bornaviridae and Filoviridae (Figure 5A. Aspiviridae, 5B. Aspiviridae). The RdRP also contains the SDD amino acid sequence in motif C, a signature for segmented negative-stranded RNA viruses (family Orthomyxoviridae and order Bunyavirales). However, phylogenetic reconstructions using sequences of the core module of RdRPs of representative negative-stranded RNA viruses reinforce the phylogenetic relatedness of the aspiviruses and suggest their separation as a monophyletic group (Figure 5A. Aspiviridae, 5B. Aspiviridae).
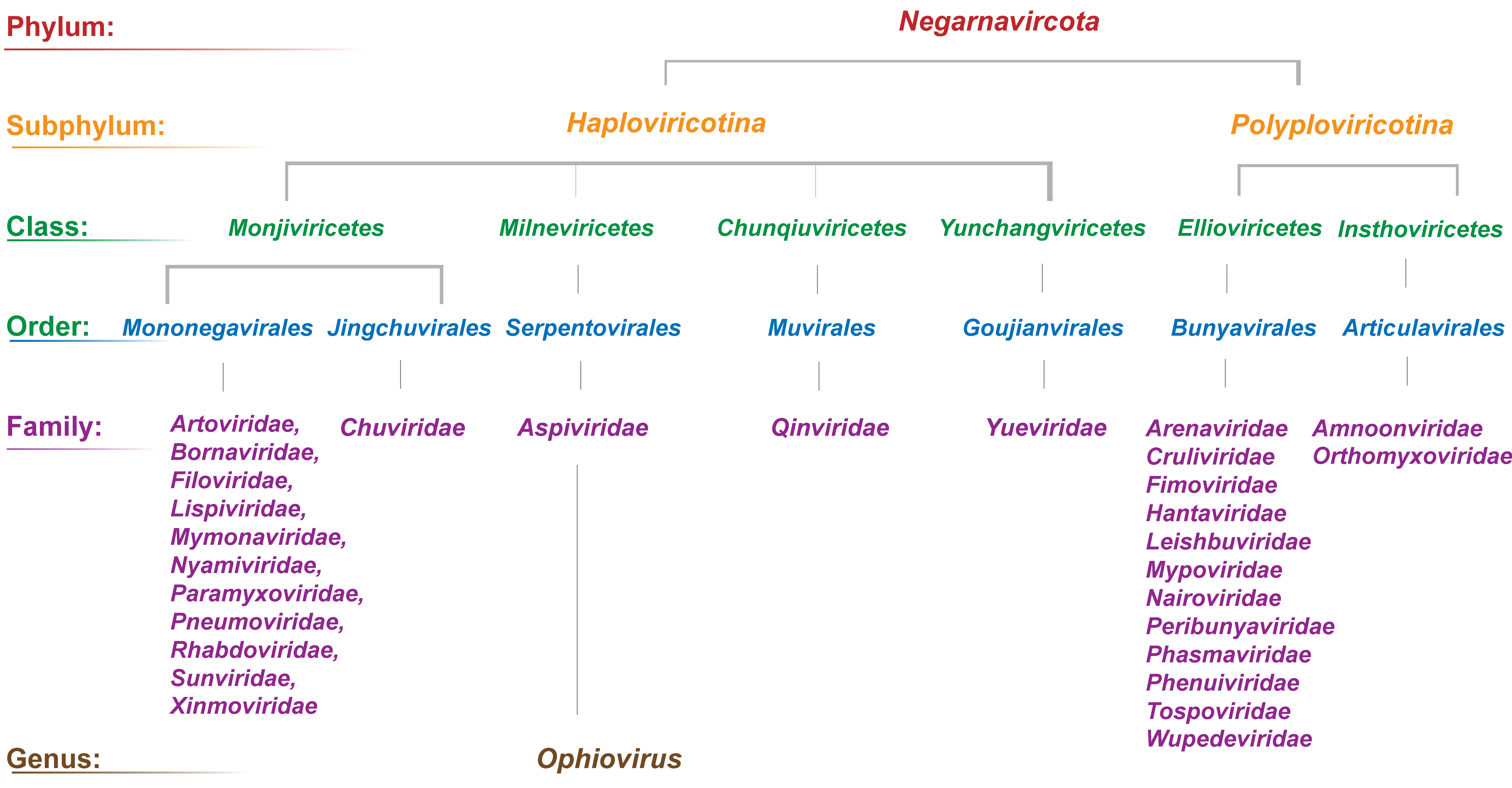 |
| Figure 4. Aspiviridae. Higher taxonomic structure of the phylum Negarnaviricota. For simplicity, only the genus Ophiovirus is indicated. |
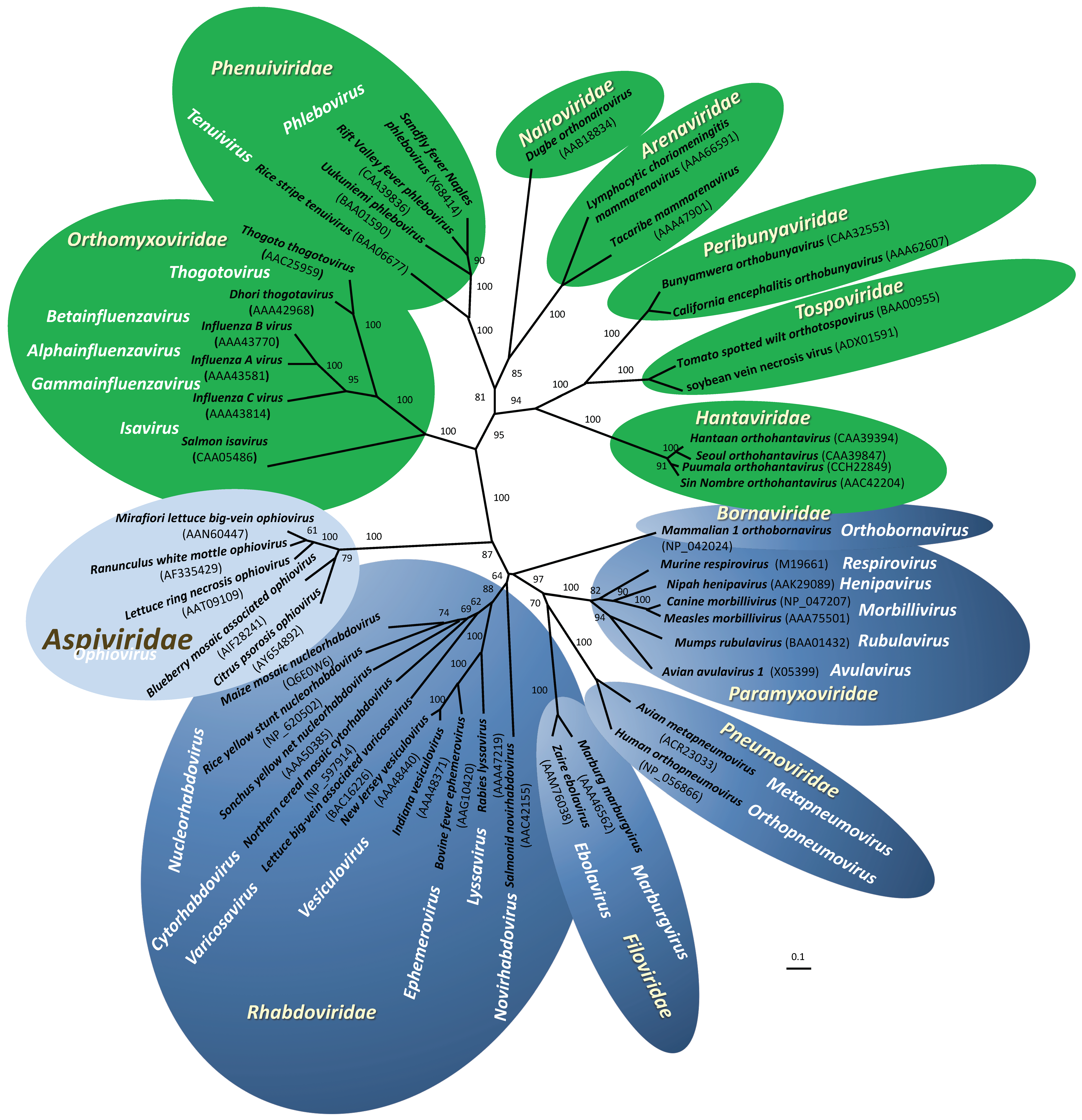 |
| Figure 5A. Aspiviridae. Phylogenetic analysis of members of the family Aspiviridae and other negative-stranded RNA viruses based on the amino acid sequences of their conserved RdRP core modules. Unrooted tree generated by the neighbor-joining method and bootstrap values (indicated for each branch node when >50%) were estimated using 100 replicates. Families are coloured according to subphyum as follows: Haploviricotina - blue (Aspiviridae - light blue), Polyploviricotina - green. This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. |
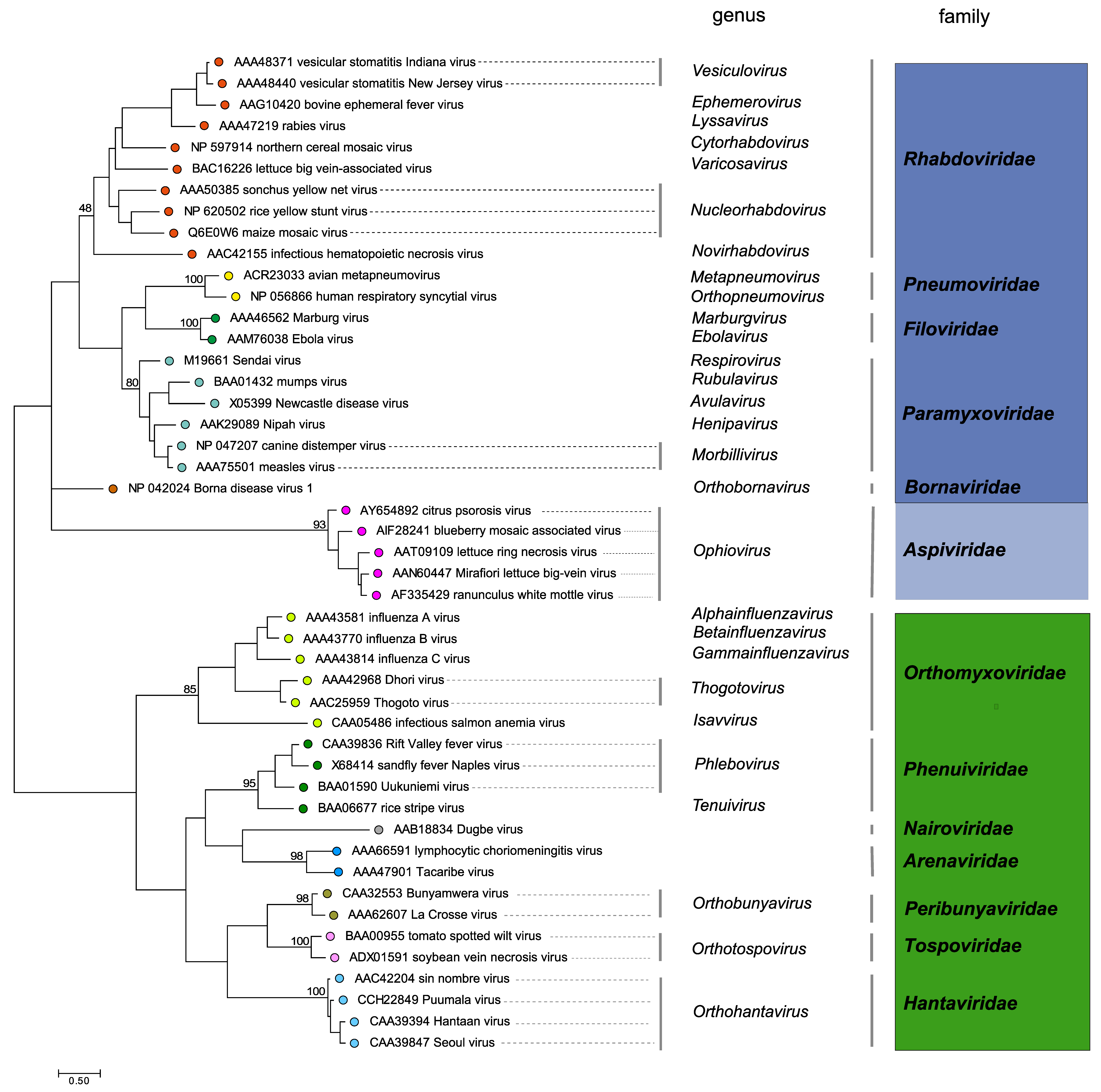 |
| Figure 5B. Aspiviridae. Phylogenetic analysis of members of the family Aspiviridae and other negative-stranded RNA viruses based on the amino acid sequences of their conserved RdRP core modules. Midpoint rooted tree generated by the neighbor-joining method and bootstrap values (indicated for each branch node when >50%) were estimated using 100 replicates. Families are coloured according to subphyum as follows: Haploviricotina - blue (Aspiviridae - light blue), Polyploviricotina - green. This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. |
Related, unclassified viruses
There are two reports regarding aspivirus-like isolates. In one study, RdRP sequences corresponding to 3 negative-stranded RNA viruses (Rhizoctonia solani negative-stranded RNA viruses 1 to 3, RsNSRV-1, -2, and -3 ) were found in the transcriptome of Rhizoctonia solani (Marzano et al., 2016) . The predicted amino acid sequences of RsNSRV-1, -2, and -3 were similar to those of the RdRP encoded by the RNA1 of LRNV and other members of the family Aspiviridae. The second report describes the detection of an RdRP sequence (Fusarium poae negative-stranded RNA virus (FpNSV) 1) in metagenomic data from Fusarium poae with 23% identity to the RdRP of MLBVV (Osaki et al., 2016). Further characterization, including particle morphology, will be required in order to define if these (putative) aspivirus-like viruses belong to the family Aspiviridae; if so they are unlikely to belong to the genus Ophiovirus.
| Virus name | Accession number | Virus abbreviation |
| Rhizoctonia solani negative-stranded RNA virus-1 | KP900919 | RsNSRV-1 |
| Rhizoctonia solani negative-stranded RNA virus-2 | KP900920 | RsNSRV-2 |
| Rhizoctonia solani negative-stranded RNA virus-3 | KP900903 | RsNSRV-3 |
| Fusarium poae negative-stranded RNA virus-1 | LC150618 | FpNSV1 |
Virus names and virus abbreviations are not official ICTV designations.