

Family: Potyviridae
Alice Kazuko Inoue-Nagata, Ramon Jordan, Jan Kreuze, Fan Li, Juan José López-Moya, Kristiina Mäkinen, Kazusato Ohshima, and Stephen J Wylie
The citation for this ICTV Report chapter is the summary published as Inoue-Nagata et al. (2022):
ICTV Virus Taxonomy Profile: Potyviridae 2022, Journal of General Virology, (2022) 103:001738.
Corresponding author: Alice Kazuko Inoue-Nagata ([email protected])
Edited by: Hélène Sanfaçon, Michael J. Adams and Luisa Rubino
Posted: March 2017, updated October 2018, February 2022, May 2023, February 2026
Summary
Members of the family Potyviridae (termed potyvirids) consist of monopartite and bipartite plant viruses with a single-stranded, positive-sense RNA genome and flexuous, filamentous particles (Table 1 Potyviridae). Genomes have a VPg covalently linked to the 5′-end, and, for most of them, the 3′-terminus is polyadenylated. Genomes encode a large polyprotein that is self-cleaved into a set of functional proteins, and another small protein produced from the genomic RNA as a trans-frame protein. Gene order is generally conserved throughout the family.
Table 1 Potyviridae. Characteristics of members of the family Potyviridae.
| Characteristic | Description |
| Example | potato virus Y-O (U09509), species Potyvirus yituberosi |
| Virion | Non-enveloped, flexuous and filamentous capsid, 650–950 nm long and 11–20 nm in diameter with a single core capsid protein |
| Genome | 8.2–11.5 kb of positive-sense, single-stranded, usually monopartite RNA (bipartite in genus Bymovirus) |
| Replication | Cytoplamsic replication in viral replication complexes residing in membranous 6K2-induced ER-derived vesicles |
| Translation | Directly from genomic RNA |
| Host range | Plants; transmission by arthropods, plasmodiophorids (Bymovirus), seeds or pollen |
| Taxonomy | Realm Riboviria, kingdom Orthornavirae, phylum Pisuviricota, class Stelpaviricetes, order Patatavirales; 13 genera including 259 species |
Virion
Morphology
Virions are flexuous filaments with no envelope and are 11–20 nm in diameter, with a helical pitch of about 3.4–3.5 nm (Figure 1 Potyviridae). Particle lengths of members of some of the 13 genera differ. Members of the genera Arepavirus, Bevemovirus, Brambyvirus, Celavirus, Ipomovirus, Macluravirus, Poacevirus, Phragmivirus, Potyvirus, Roymovirus, Rymovirus and Tritimovirus are monopartite with particle modal lengths of 650–950 nm. Members of the genus Bymovirus are bipartite with particles of two modal lengths of 250–300 nm and 500–600 nm.
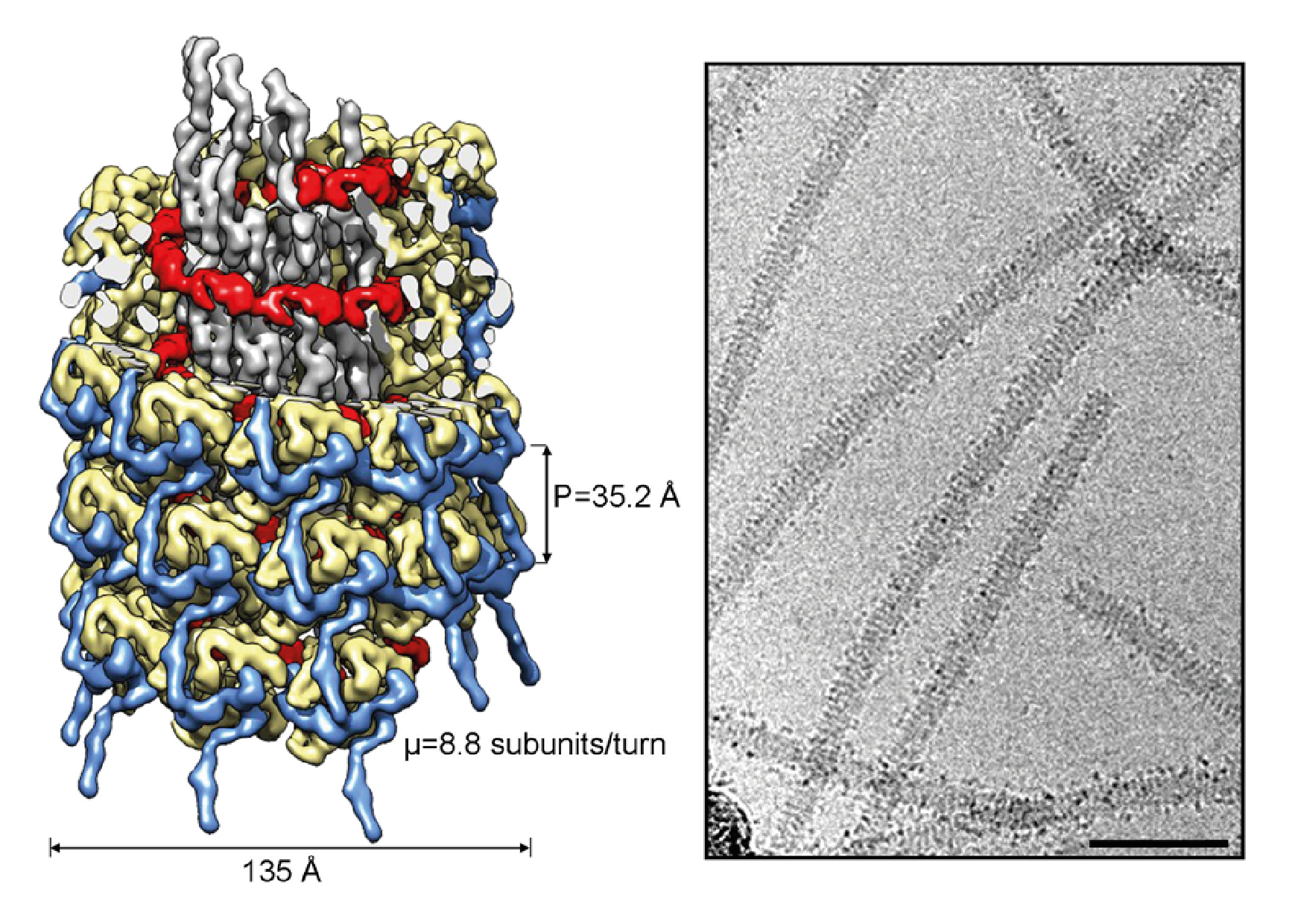 |
| Figure 1 Potyviridae (Left) Rendering of the cryoEM map for a section of a virion of turnip mosaic virus (TuMV), using color codes for the ssRNA (red) and for the different regions of the coat proteins: central globular domain (pale yellow), N-terminal arm (blue), and C-terminal region (grey). In the upper section, some of the densities are cut away to visualize better the ssRNA and the C-terminal regions of the coat proteins that construct the inner side of the helical virus. The first 60 amino acids at the N-terminus are exposed to the solvent and are not seen in the cryoEM map since they are flexible. Calculated dimensions for width, pitch, and number of subunits per turn are indicated. (Right) Representative cryoEM micrograph of TuMV virions where the periodic organization of the helical filaments is apparent. Scattered globular densities surrounding the filaments suggest the presence of local folded domains in the N-terminal region that is missing in the cryoEM map. Bar equals approximately 500 nm. Images are courtesy of R. Cuesta and M. Valle, CIC bioGUNE, Spain, based on (Cuesta et al., 2019). |
Physicochemical and physical properties
Virions of viruses in the genera Potyvirus and Rymovirus have a density in CsCl of about 1.31 g cm−3 and sedimentation coefficient S20,w of 137–160S. Those of the genus Bymovirus have a density in CsCl of about 1.29 g cm−3.
Nucleic acid
Viruses in all genera, except Bymovirus, have a single molecule of positive-sense, single-stranded (ss) RNA of 8.2 kb (bellflower veinal mottle virus-SW, genus Bevemovirus; artichoke latent virus-FR37, FR50, genus Macluravirus) to 11.5 kb (celery latent virus, genus Celavirus). Virions are infectious. A tract of 20 to 160 adenosine residues is present at the 3′-terminus, except for the celavirus celery latent virus. Bymoviruses have two positive-sense, ssRNA molecules; RNA1 is 7.3–7.6 kb and RNA2 is 3.5–3.7 kb. Both RNAs have 3′-terminal polyadenylate tracts and probably a VPg at their 5′-termini.
Proteins
Virions contain one type of coat (capsid) protein (CP) of 28.5–47 kDa. N-terminal residues are positioned on the exterior of the virion (Figure 1.Potyviridae) (Zamora et al., 2017, Cuesta et al., 2019). Mild trypsin treatment removes N- and C-terminal segments, leaving a trypsin-resistant core of about 24 kDa. Plant proteases may degrade the CP in vivo, as occurs in vitro during purification using some procedures or from particular hosts. All potyviral CPs display significant aa sequence identity in the trypsin-resistant core, but little identity in their N and C-terminal segments. A virus-encoded viral protein genome-linked (VPg) of about 24 kDa is covalently linked to the 5′-terminal nucleotide. The particles may also contain other virus-encoded proteins like the helper component protease (HC-Pro) (Torrance et al., 2006), or the cylindrical inclusion (CI) protein (Gabrenaite-Verkhovskaya et al., 2008), and the host factor elF4E (Tavert-Roudet et al., 2017).
Genome organization and replication
Genomic RNA (two RNAs for members of the genus Bymovirus) encodes a single major polyprotein (Figure 2 Potyviridae). Typically, this then undergoes co- and post-translational proteolytic processing by two or three virus-encoded proteinases to form the mature proteins. Genomic RNA replicates via the production of a full-length negative-sense RNA. While there are exceptions noted in the relevant genus descriptions, the polyprotein of most monopartite viruses in the family is cleaved into ten products, which show conservation of sequence and organization (Adams et al., 2005a, Yang et al., 2021). A second small ORF, PIPO, is generated by a polymerase slippage mechanism and is expressed as the trans-frame protein P3N-PIPO (Chung et al., 2008, Rodamilans et al., 2015). Another additional small ORF, PISPO, is generated through a transcriptional slippage mechanism among sweet potato-subgroup potyviruses and leads to the production of a trans-frame protein, P1N-PISPO (Li et al., 2012, Mingot et al., 2016, Olspert et al., 2016, Untiveros et al., 2016). Potyvirus protein products are listed below with their most important functions (details from (Revers and García 2015, Yang et al., 2021)):
- P1 (protein 1 protease): Of all the potyvirid proteins, P1 is the least conserved in sequence and the most variable in size. It plays a significant role in virus replication, probably due to the stimulation of the gene silencing suppressor HC-Pro and/or suppression of host immune responses. A serine protease domain towards the C-terminus cleaves P1 from the polyprotein in a host factor independent-manner, typically at Tyr/Phe-Ser.
- HC-Pro (helper component-protease): The HC-Pro protein has roles in suppression of gene silencing and in vector transmission. A cysteine protease domain towards the C-terminus cleaves it from the remainder of the downstream polyprotein, typically at Gly-Gly. It is also an important factor for the formation of stable virions.
- P3 (protein 3): Involved in virus replication, movement and appears to be significant in host range and symptom development.
- 6K1 (6-kDa peptide 1): Although it is present in the replication complex and required for replication, the function of this small protein is not known.
- CI (cylindrical inclusion protein): This protein has helicase activity and accumulates in inclusion bodies in the cytoplasm and plasmodesmata of infected plant cells.
- 6K2 (6-kDa peptide 2): A small transmembrane protein probably anchoring the replication complex to the ER.
- VPg (viral protein genome-linked): The first region of the NIa protein, susceptible to cleavage and sepatation from the protease region (Pro). Attached to the 5′-terminus of the genome and belongs to a class of intrinsically disordered proteins. It is essential for virus replication and translation, interacting with one or several isoforms of the eIF4E translation initiation factor, and is also involved in suppression of RNA silencing. Accumulates as inclusions in the nucleus.
- NIa-Pro (nuclear inclusion a-protease): The second region of the NIa protein. Serine-like cysteine protease responsible for cleavage of most sites in the polyprotein, typically at Gln/Glu-Ser/Gly/Ala. Accumulates as inclusions in the nucleus.
- NIb (nuclear inclusion b): The RNA-directed RNA polymerase; also involved in suppressing host defenses. Accumulates as inclusions in the nucleusi.
- CP (coat (capsid) protein): Viral coat protein that also has roles in virus movement, genome amplification and vector transmission.
- PIPO (pretty interesting Potyviridae ORF): A short ORF embedded within the P3 cistron expressed as the trans-frame P3N-PIPO protein by a polymerase slippage mechanism; identified throughout the family and shown to be essential for the intercellular movement of viruses.
- PISPO (pretty interesting sweet potato potyvirus ORF): Some sweet potato potyviruses encode an additional ORF, embedded within the C-terminal part of an exceptionally large P1 protein. A transcriptional slippage mechanism leads to production of a trans-frame protein, P1N-PISPO, that functions as an RNA silencing suppressor.
The polymerase slippage model for the production of P3N-PIPO is consistent with the theory that potyvirus intercellular movement is coupled to active virus genome replication and translation. The genomic RNA replicates via the production of a full-length negative-sense RNA, which takes place in the viral replication complex (VRC) in the cytoplasm. The VRC is formed by virus proteins such as NIb, 6K2, VPg, NIa-Pro, HC-Pro, and CI, and possiby also P3 and 6K1 (Cui and Wang 2016, Cui et al., 2017), as well as various host factors. Potyvirus replication initiates at 6K2-induced ER-originated vesicles. The cellular COPI and COPII coating machineries are involved in the biogenesis of potyvirus 6K2 vesicles (Wei and Wang 2008). The ER-derived 6K2 vesicles also target and are associated with chloroplasts for virus replication. The viral genome is transported from the cytoplasm through the plasmodesmata, most likely in the form of intact particles, and involves the coordinated action of CP, CI, P3, 6K2, and P3N-PIPO (Chai et al., 2020, Wang 2021).
 |
| Figure 2 Potyviridae. Genome organization of representatives of Potyviridae genera. The ssRNA genome is represented by a line and the polyprotein ORF by an open box with the mature proteolytic products named. P1 (protein 1 protease), a protein with serine proteolytic activity responsible for cleavage at typically Tyr/Phe-Ser; HC-Pro (helper component protease), a protein with aphid transmission helper-component activity and cysteine proteolytic activity responsible for cleavage at typically Gly-Gly; P3 (protein 3); PIPO (pretty interesting Potyviridae ORF; expressed as the trans-frame P3N-PIPO protein); 6K1 (6 kDa peptide 1); CI (cylindrical inclusion); 6K2 (6 kDa peptide 2); VPg (virus protein genome-linked), the genome-linked virus protein covalently attached to the 5 -terminal nucleotide; represented here by an oval; NIa-Pro (the second of two functional regions of the NIa, nuclear inclusion a protease, susceptible to being proteolytically separated from the VPg region), a cysteine-like proteolytic activity responsible for cleavage at Gln/Glu-(Ser/Gly/Ala); NIb (nuclear inclusion b), RNA-directed RNA polymerase; CP (coat protein). The P1 cistron of the brambyvirus genome contains an embedded AlkB domain. Cleavage sites of P1 (blue arrow), HC-Pro (red arrow), and NIa-Pro (black arrow) are indicated. |
Biology
Inclusion body formation
All members of the family Potyviridae form cytoplasmic cylindrical inclusion (CI) bodies during infection. The CI is an array of a 70 kDa viral protein that possesses ATPase and helicase activities. Some potyviruses induce nuclear inclusion bodies that are co-crystals of two viral-encoded proteins – NIa and NIb – that are present in equimolar amounts. The small nuclear inclusion (NIa) protein (49 kDa) is a polyprotein consisting of the VPg and NIa-Pro domains. The large nuclear inclusion (NIb; 60 kDa) contains the RNA-directed RNA polymerase domain responsible for viral genome replication. Amorphous inclusion bodies (potyvirus-induced granules) are also evident in the cytoplasm during certain potyvirus infections and represent aggregations of HC-Pro (Hafrén et al., 2015) and perhaps other non-structural proteins.
Host range
Most members have a narrow natural host range, but can infect a wider range of experimental hosts, and a few members infect species in up to 30 plant families. Transmission to experimental hosts is readily accomplished by mechanical inoculation. Many viruses are widely distributed. Distribution is aided by seed transmission in some cases.
Transmission
Potyvirids are vectored by a variety of organisms. Members of the genera Potyvirus and Macluravirus have aphid vectors that transmit in a non-persistent, non-circulative manner. Two helper component protease (HC-Pro) aa motifs (R/KITC and PTK) and a CP motif (DAG for many potyviruses) are highly conserved and are required for aphid transmission. The two known arepaviruses have two HC-Pro-like cistrons, but the role of the encoded proteins in transmission, and the identity of the virus vectors are unknown. Viruses in other genera in the family Potyviridae lacking these motifs are not transmitted by aphids, and include rymoviruses, poaceviruses and tritimoviruses which are transmitted by eriophyid mites in a semi-persistent manner. The vector of rose yellow mosaic virus (RoYMV, species Roymovirus rosae) has not been identified. RoYMV lacks the conserved potyvirus HC-Pro motifs and instead, a putative C-2x-C eriophyid mite transmission motif occurs at the N-terminus of HC-Pro. The DAG motif of CP is also lacking. Together this information suggests RoYMV may also be transmitted by eriophyid mites, although this has not been proven experimentally. Bymoviruses are transmitted by root-infecting plasmodiophorids. Ipomoviruses appear to be transmitted by whiteflies. The vector of members of the single species in the genera Brambyvirus and Celavirus are unknown. The vector of bellflower veinal mottle virus (BVMV), a member of the only species in the genus Bevemovirus, has not been identified. BVMV HC-Pro lacks the conserved potyvirus aphid transmission motifs, but its CP has a DTG motif near its N-terminus, possibly analogous to the DAG motif involved in aphid transmission. The vector responsible for the transmission of the two phragmiviruses remains unknown. It is believed that they are not transmitted by aphids, as the three conserved aphid-transmission motifs typically found in the HC-Pro (KITC- and PTK-like motifs) and in the CP (DAG motif) are absent from the phragmivirus genome.
Potyvirids can be mechanically transmitted to other hosts in experimental studies. Contact transmission of some potyviruses between adjacent plants may occur naturally (Jones 2018). Some viruses are seed�borne, and some are pollen transmitted, such as zucchini yellow mosaic virus (Harth et al., 2017).
Antigenicity
An epitope of CP in the conserved internal trypsin-resistant core has been identified that is similar in most members of the family. Virions are moderately immunogenic; there are serological relationships among many members.
Derivation of names
Arepavirus: from areca palm necrotic spindle-spot virus, a member of the species Arepavirus arecamaculatum, that produces symptoms upon infection of Dypsis lutescens (areca palm)
Bevemovirus: from symptoms (veinal mottle) induced on Campanula takesimana (bellflower) by bellflower veinal mottle virus, a member of the species Bevemovirus campanulae
Brambyvirus: from bramble, the common name for several Rubus species, some of which are infected by blackberry virus Y, a member of the species Brambyvirus rubi
Bymovirus: from symptoms (yellow mosaic) on barley leaves induced by barley yellow mosaic virus, a member of the species Bymovirus hordeiluteum
Celavirus: from celery latent virus, a member of the species Celavirus apii responsible for the apparently symptomless infection of celery
Ipomovirus: from Ipomea batatas (sweet potato) and mosaic, the symptoms induced on it by infection of sweet potato mild mottle virus, a member of the species Ipomovirus lenisbatatae
Macluravirus: from the genus name of the host, Maclura pomifera, of Maclura mosaic virus, a member of the species Macluravirus maclurae
Phragmivirus: from Phragmites australis, the host of common reed chlorotic stripe virus, a member of the species Phragmivirus phragmii
Poacevirus: from Poaceae, the family name of the grass hosts of Triticum mosaic virus, a member of the species Poacevirus tritici
Potyviridae, Potyvirus: from potato virus Y, a member of the species Potyvirus yituberosi
Roymovirus: from a description of the symptoms (yellow mosaic) induced on Rosa host species by rose yellow mosaic virus, a member of the species Roymovirus rosae
Rymovirus: from a description of the symptoms (mosaic) induced on ryegrass (Lolium spp.) by ryegrass mosaic virus, a member of the species Rymovirus lolii
Tritimovirus: from the host genus (Triticum) and induced symptoms (mosaic) by isolates of wheat streak mosaic virus, a member of the species Tritimovirus tritici
Genus demarcation criteria
The 13 genera are differentiated by biological criteria, mainly transmission by specific vectors, and by molecular data (Adams et al., 2005b, Gibbs and Ohshima 2010, Gibbs et al., 2020, Moury and Desbiez 2020). Over the entire open reading frame, members of different genera are <46% identical in nucleotide sequence, with the exception of the genera Rymovirus and Potyvirus, members of which have different vectors.
Species demarcation criteria
Members of different species have complete ORF sequences that are generally <76% identical in nucleotide sequence and <82% identical in amino acid sequence. The thresholds for species demarcation using nucleotide identity values for the individual coding regions range from <58% for the P1 coding region to <74–78% for other regions. For the coat protein, the optimal species demarcation criterion is <76–77% nucleotide and <80% amino acid identity (Shukla and Ward 1989, Adams et al., 2005b). In considering the evidence for new species or genera in the family Potyviridae, the Study Group will evaluate each new case based on complete or near-complete genome sequence(s) together with host and biological characteristics.
Relationships within the family
Phylogenetic analysis of complete polyprotein sequences of viruses in the family Potyviridae reveals clades corresponding to the 13 genera (Figure 3 Potyviridae).
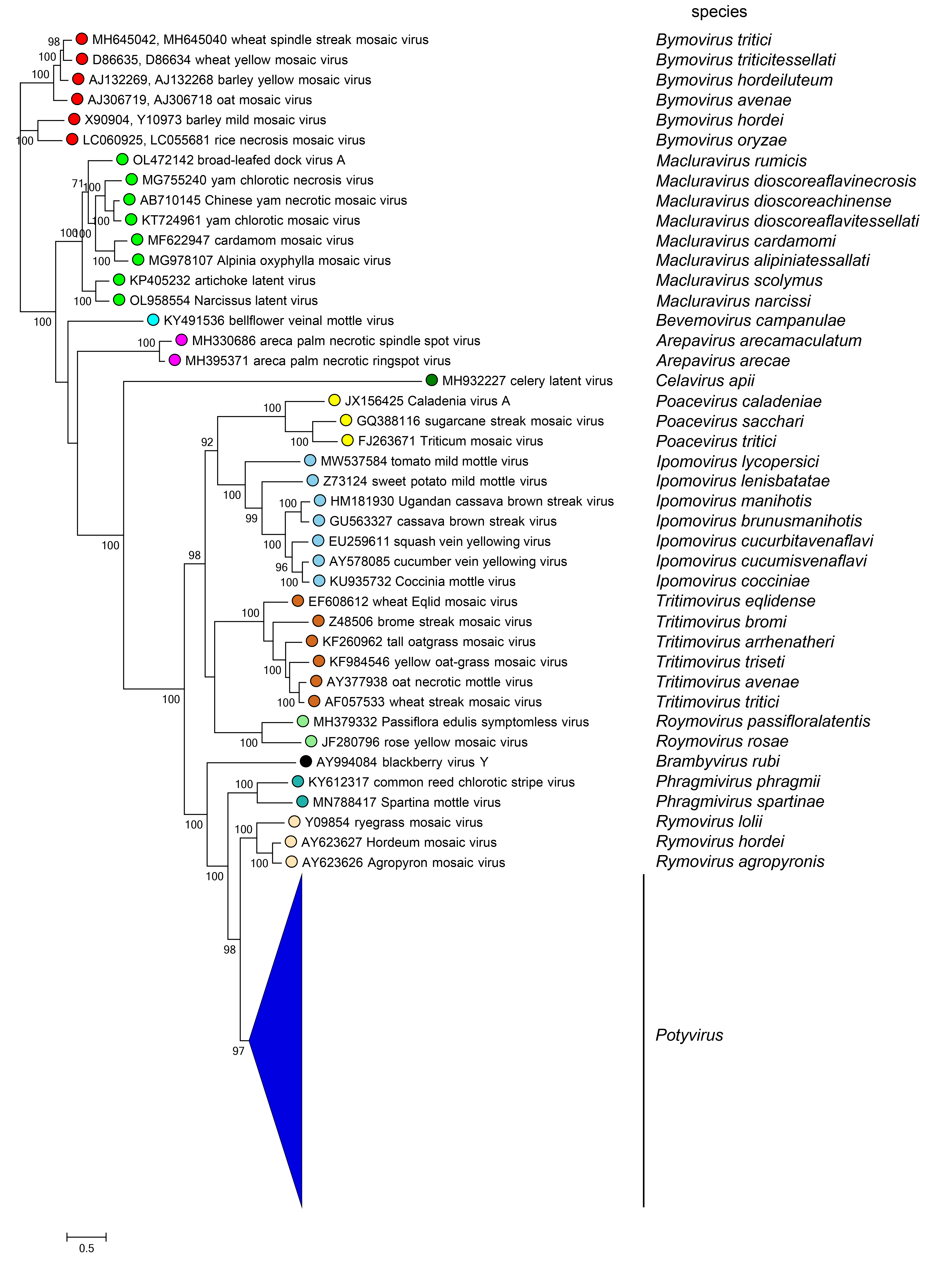 |
| Figure 3 Potyviridae. Phylogenetic tree contructed using the complete polyprotein amino acid sequences of representative viruses within the family Potyviridae. The maximum likelihood (ML) tree was produced in PhyML v.3.1 (Guindon et al., 2010) using the LG model (Le and Gascuel 2008) with gamma-distributed among-site rate variation; and a proportion of invariable sites was selected as the best substitution model by ModelTest-NG (Darriba et al., 2020) from a degapped CLUSTAL X version 2 (Larkin et al., 2007) alignment of polyprotein amino acid sequences, implemented in FigTree (http://tree.bio.ed.ac.uk/software/Figtree/). Bootstrap values > 70% are given at nodes. The scale bar represents 0.5 substitutions per site. The midpoint rooted ML tree was visualized using FigTree v1.4.4 (http://tree.bio.ed.ac.uk/software/figtree/).This phylogenetic tree and corresponding sequence alignment are available to download from the Resources page. Tree constructed by Kazusato Ohshima. A full version of this tree showing all sequences is provided on a separate page. |
Relationships with other taxa
Members of the family Potyviridae are related to viruses in the order Picornavirales in their genome structure, in particular, the presence of a single ORF translated into a polyprotein and the block of replication genes (helicase, VPg, 3C-like protease, and polymerase). However, they differ from picornaviruses in virion morphology, the size of VPg, and the type of helicase.