

Family: Bromoviridae
Jeremy R. Thompson, Tomás Canto, John P. Carr, Vicente Pallás and Dana Šafářová.
The citation for this ICTV Report chapter is the summary published as Bujarski et al., (2019):
ICTV Virus Taxonomy Profile: Bromoviridae, Journal of General Virology, 100, 1206–1207.
Corresponding author: Jeremy R. Thompson (Jeremy.thompson@mpi.govt.nz)
Edited by: F. Murilo Zerbini, Sead Sabanadzovic and Luisa Rubino
Posted: May 2019, updated September 2022
PDF: ICTV_Bromoviridae.pdf (2019 version)
Summary
The family Bromoviridae includes plant viruses with tri-segmented, positive-sense RNA genomes of about 8 kb. Genomic RNAs are packaged in separate virions that may also contain subgenomic, defective or satellite RNAs. Virions are variable in morphology (spherical or bacilliform) and are transmitted mechanically, in/on pollen, and non-persistently by insect vectors. Members of the family cause major disease epidemics in fruit, vegetable and fodder crops such as tomato, cucurbits, bananas, and alfalfa.
Table 1.Bromoviridae. Characteristics of members of the family Bromoviridae
| Characteristic | Description |
| Example | brome mosaic virus (RNA1: X02380; RNA2: X01678; RNA3: J02042), species Bromovirus BMV |
| Virion | Spherical or quasi-spherical (26–35 nm diameter) or bacilliform (18–26 nm diameter, 30–85 nm long) |
| Genome | Three segments of linear positive-sense single-stranded RNA, comprising about 8 kb in total |
| Replication | On cytoplasmic membranes with genomic RNAs. Coat protein may be required for genome activation |
| Translation | Directly from genomic or subgenomic RNAs |
| Host range | From narrow to broad range of plants |
| Taxonomy | Realm Riboviria, kingdom Orthornavirae, phylum Kitrinoviricota, class Alsuviricetes, order Martellivirales; six genera, including 52 species |
Virion
Morphology
Virions are either spherical or quasi-spherical (Figure 1.Bromoviridae, panels A–D), having T=3 icosahedral symmetry, and a diameter of 26–35 nm (members of the genera Anulavirus, Bromovirus, Cucumovirus and Ilarvirus) or bacilliform (members of the genera Alfamovirus, Ilarvirus and Oleavirus) having (within members of a species) diameters of 18–26 nm and lengths of 30–85 nm, depending on the virus. Genomic RNAs are packaged in separate virions that may also contain subgenomic RNAs (sgRNAs), defective RNAs or satellite RNAs (Table 2.Bromoviridae).
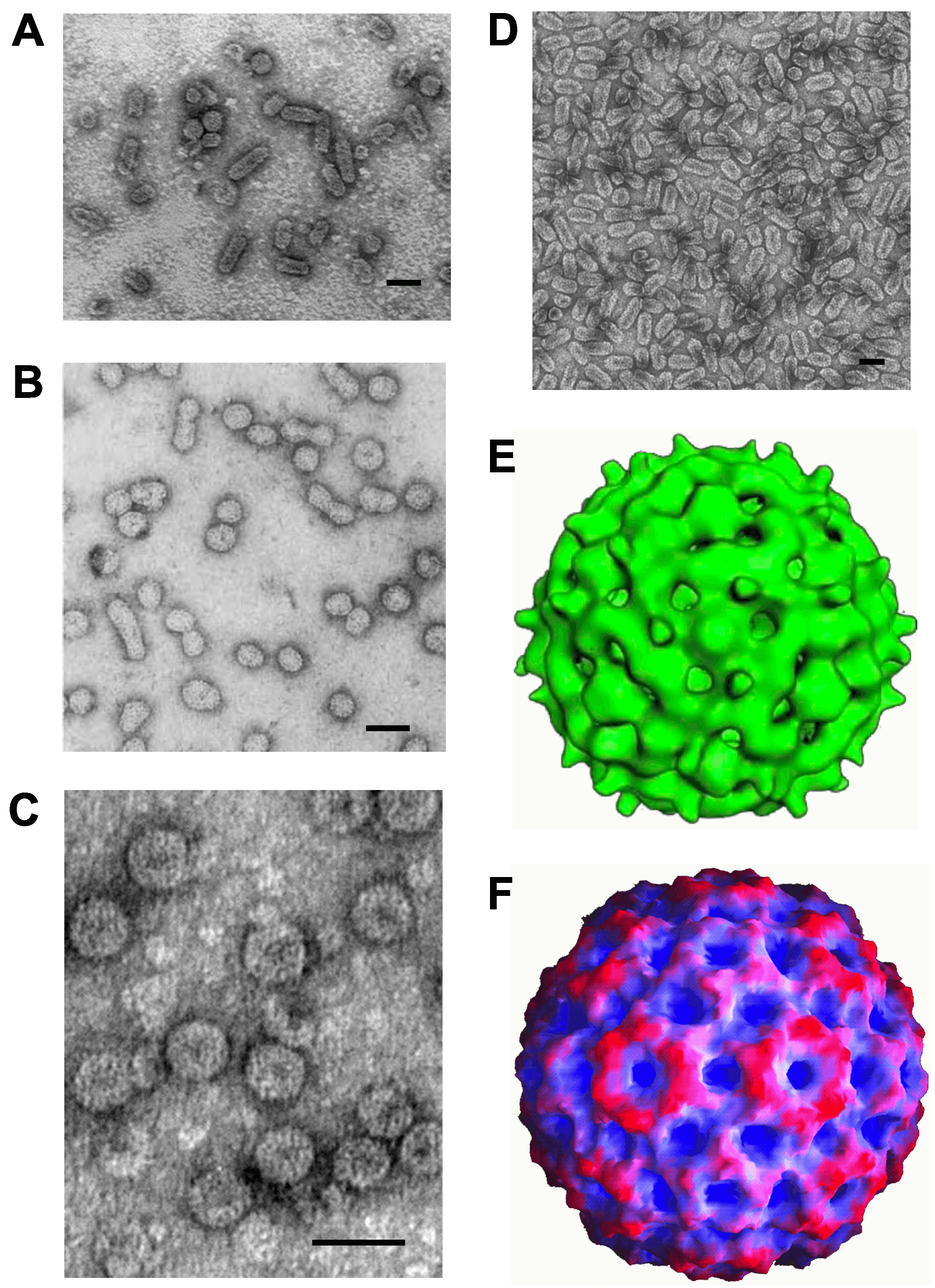 |
| Figure 1.Bromoviridae. Negative-contrast electron micrograph of particles of (A) alfalfa mosaic virus, (B) prunus necrotic ringspot virus, (C) cucumber mosaic virus and (D) olive latent virus 2. Reconstruction of particles of (E) tobacco streak virus and (F) cucumber mosaic virus. Image B reprinted from (Pallas et al., 2013), with permission; image F, courtesy of Dr. K.L. Perry, Cornell University, Ithaca, NY, USA and Dr. T. Smith, University of Texas, Galveston, Texas, USA; Images A and C–E, courtesy of A. De Stradis, IPSP-CNR, Bari, Italy; images B and F courtesy of A. Paredes, NCTR/ORA, Arkansas USA. Bars = 50 nm. |
Physicochemical and physical properties
The Mr of virions varies from 3.5 to 6.9×106, depending on the nucleic acid content. The buoyant density of formaldehyde-fixed virions ranges from 1.35 to 1.37 g cm−3 in CsCl. The S20,w varies from 63S to 99S. Virion integrity is dependent on RNA–protein interactions and virion RNA is susceptible to RNase degradation in situ at neutral pH. Heat inactivation occurs at 60 °C for members of some genera; this has not been tested for all genera. In some cases, virions are unstable in the presence of divalent cations. Virions are generally stable in the presence of chloroform, but not in the presence of phenol. Virions are unstable in the presence of strong anionic detergents such as SDS but can tolerate low concentrations of mild detergents such as Triton X-100.
Nucleic acid
Total genome length is approximately 8 kb. Genomes consist of three linear, positive-sense single-stranded (ss) RNAs with 5′-terminal cap structures. The 3′-termini are not polyadenylated, but generally are highly conserved within members of the same species or isolate, and form strong secondary structures. They are either tRNA-like and can be aminoacylated (members of the genera Bromovirus and Cucumovirus) or form other structures that are not aminoacylated (members of the genera Alfamovirus, Anulavirus, Ilarvirus and Oleavirus) (Table 2.Bromoviridae).
Table 2.Bromoviridae Genome characteristics of viruses belonging to a representative species of each genus in the family Bromoviridae
| ||||||
Genus Species | Isolate | RNA1 (nt) | RNA2 (nt) | RNA3 (nt) | 3' terminus | DI / sat RNAs |
Alfamovirus Alfalfa mosaic virus |
425 |
3644 |
2593 |
2037 |
Complex* |
−/− |
Anulavirus Pelargonium zonate spot virus |
Tomato |
3383 |
2435 |
2659 |
Complex* |
−/− |
Bromovirus Brome mosaic virus |
Russian |
3234 |
2865 |
2117 |
tRNA† |
+/− |
Cucumovirus Cucumber mosaic virus |
FNY |
3357 |
3050 |
2216 |
tRNA† |
+/+ |
Ilarvirus Tobacco streak virus |
WC |
3491 |
2926 |
2205 |
Complex* |
−/− |
Oleavirus Olive latent virus 2 |
Apulia |
3126 |
2734 |
2438 |
Complex* |
? |
DI / sat: presence of defective interfering / satellite RNAs
* Complex secondary structure.
† Aminoacylatable, with pseudoknot folding.
Proteins
A single 20–24 kDa coat protein (CP) is expressed from a sgRNA. The CP is generally required for systemic movement and may be required for cell-to-cell spread in some cases. CP may also be required for virus expression and replication (members of the genus Alfamovirus) or activation of replication (members of the genus Ilarvirus). The phenomenon of genome activation has been demonstrated to cross group boundaries since the CP of an alfamovirus can activate the genome of an ilarvirus and vice versa. An intriguing feature of the alfamovirus CP is its accumulation in nucleus and nucleolus (Herranz et al., 2012).
Lipids
Lipids are not associated with virions.
Carbohydrates
Carbohydrates are not associated with virions.
Genome organization and replication
RNA1, RNA2, and RNA3 can act as mRNAs. The genomic RNA1 and RNA2 each encodes a single large ORF. Proteins encoded by these ORFs (1a and 2a) act with host factors as the viral replicase. In members of the genus Cucumovirus and in some members of the genus Ilarvirus (subgroups 1 and 2 only) a small 2b protein is expressed from an additional sgRNA that may or may not be encapsidated. These 2b proteins are involved in cell-to-cell movement, suppression of post-transcriptional gene silencing and symptom induction. The 3a protein is a movement protein and ORF3b encodes the CP, expressed from a sgRNA (Figure 2.Bromoviridae; Table 3.Bromoviridae).
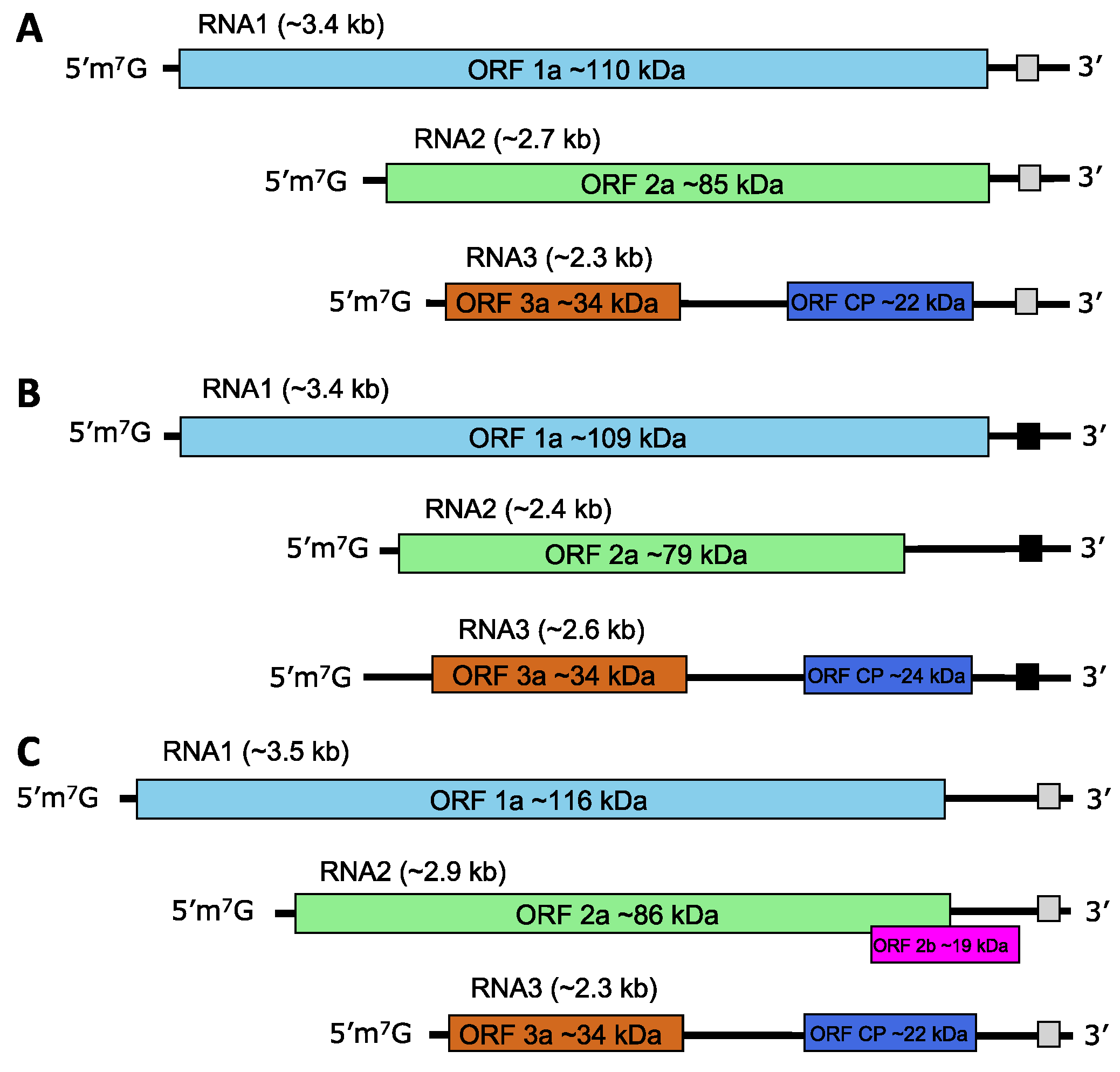 |
| Figure 2.Bromoviridae. Schematic genome organization of members of the family Bromoviridae: members of (A) the genera Alfamovirus, Bromovirus, Ilarvirus subgroups 3 and 4 and Oleavirus; (B) the genus Anulavirus; (C) the genera Cucumovirus and Ilarvirus subgroups 1 and 2. The 3′-termini form either tRNA-like (B) or complex structures (A, C) shown as black or grey squares. |
Table 3.Bromoviridae Features of the proteins encoded by members of the family Bromoviridae
| Protein | Mass (kDa) | RNA | Function* |
|---|---|---|---|
| 1a | 102.7–125.8 | RNA1 | Methyltransferase, helicase |
| 2a | 78.9–96.7 | RNA2 | RNA-directed RNA polymerase (RdRP) |
| 2b | 12.7- 21.0 | RNA2 | Cell-to-cell movement, suppression of post-transcriptional gene silencing, symptom induction |
| 3a | 30.5–36.5 | RNA3 | Cell-to-cell movement |
| CP | 19.8–26.2 | sgRNA4† | Encapsidation, movement, genome activation |
* Functions of 1a and 2a are putative in most cases, by analogy to related viruses.
† The sgRNA for the CP derived from RNA3 is encapsidated in members of all genera except Oleavirus.
There is no clear evidence of proteolytic or other post-translational processing. Virus replication occurs on cytoplasmic membranes via full-length negative-sense strand (−) synthesis and subsequent positive-sense (+) strand synthesis. The sgRNAs are synthesized from the (−) template, and may or may not be found in the virions. CP accumulates to high levels in infected cells, whereas the nonstructural proteins accumulate to much lower levels. Virions accumulate in the cytoplasm. The replication cycle of the virus takes place predominantly in the cytoplasm, although in the case of members of the genus Alfamovirus it has been demonstrated that a cytoplasmic/nuclear balance of CP accumulation accurately modulates viral expression (Pallas et al., 2013).
Biology
The natural host range of the viruses spans from very narrow (members of the genera Bromovirus, Oleavirus) to extremely broad (members of the genus Cucumovirus). Members of the family cause major disease epidemics in fruit, vegetable and fodder crops such as tomato (Figure 3.Bromoviridae), cucurbits, bananas, and alfalfa.
Bromoviruses are transmitted mechanically, in/on pollen, through seeds, and non-persistently by insect vectors.
 |
| Figure 3.Bromoviridae. Severe outbreaks of cucumber mosaic virus containing necrogenic satellite RNA (A) and pelargonium zonate spot virus (B) in crops of canning tomato in southern Italy. Insets show disease symptoms on fruits. |
Antigenicity
Native virions are generally poor immunogens and require stabilization with formaldehyde prior to use as antigens. There are few or no serological relationships between members of different genera, and weak relationships between members of the same genus.
Genus demarcation criteria
Genera in the family have been designated on the basis of natural host range, mode of transmission, particle morphology and physicochemical properties, genome structure and replication strategies, and ability to support replication of defective RNAs and satellite RNAs.
Derivation of names
Alfamovirus: from alfalfa mosaic virus.
Anulavirus: from Latin “anularis” (ring-shaped) describing the concentric symptom induced in tomato by members of the species Pelargonium zonate spot virus
Bromoviridae, Bromovirus: from Brome mosaic, also, from Bromus (genus of the host of brome mosaic virus).
Cucumovirus: from cucumber mosaic virus.
Ilarvirus: from the disease symptom isometric labile ringspot.
Oleavirus: from Olea, the genus name of olive, the host for members of the genus.
Relationships within the family
Relationships within this family have been examined using the complete genomic sequence, genes and gene products. The three genera with multiple members (Bromovirus, Cucumovirus and Ilarvirus) form distinct clades. Members of the genera Anulavirus and Oleavirus are unique and distinct from those of other genera within the family. A longstanding debate as to whether alfalfa mosaic virus (AMV, genus Alfamovirus) should be regarded as a member of the genus Ilarvirus has yet to be resolved. Ilarviruses share many features at the molecular level with AMV, including the activation of ilarvirus replication by AMV CP and vice versa. However, a major difference between AMV and ilarviruses is the mode of transmission: AMV is transmitted non-persistently by aphids, whereas ilarviruses are transmitted through pollen and feeding of thrips. Studies defining recombination events in the genomes of members of the Bromoviridae suggest that AMV should be grouped with the ilarviruses (Pallas et al., 2013). Except for AMV, members of all genera can be phylogenetically delineated using a sequence flanked by the highly conserved the RdRP motifs I-VII (Koonin 1991) (Figure 4.Bromoviridae).
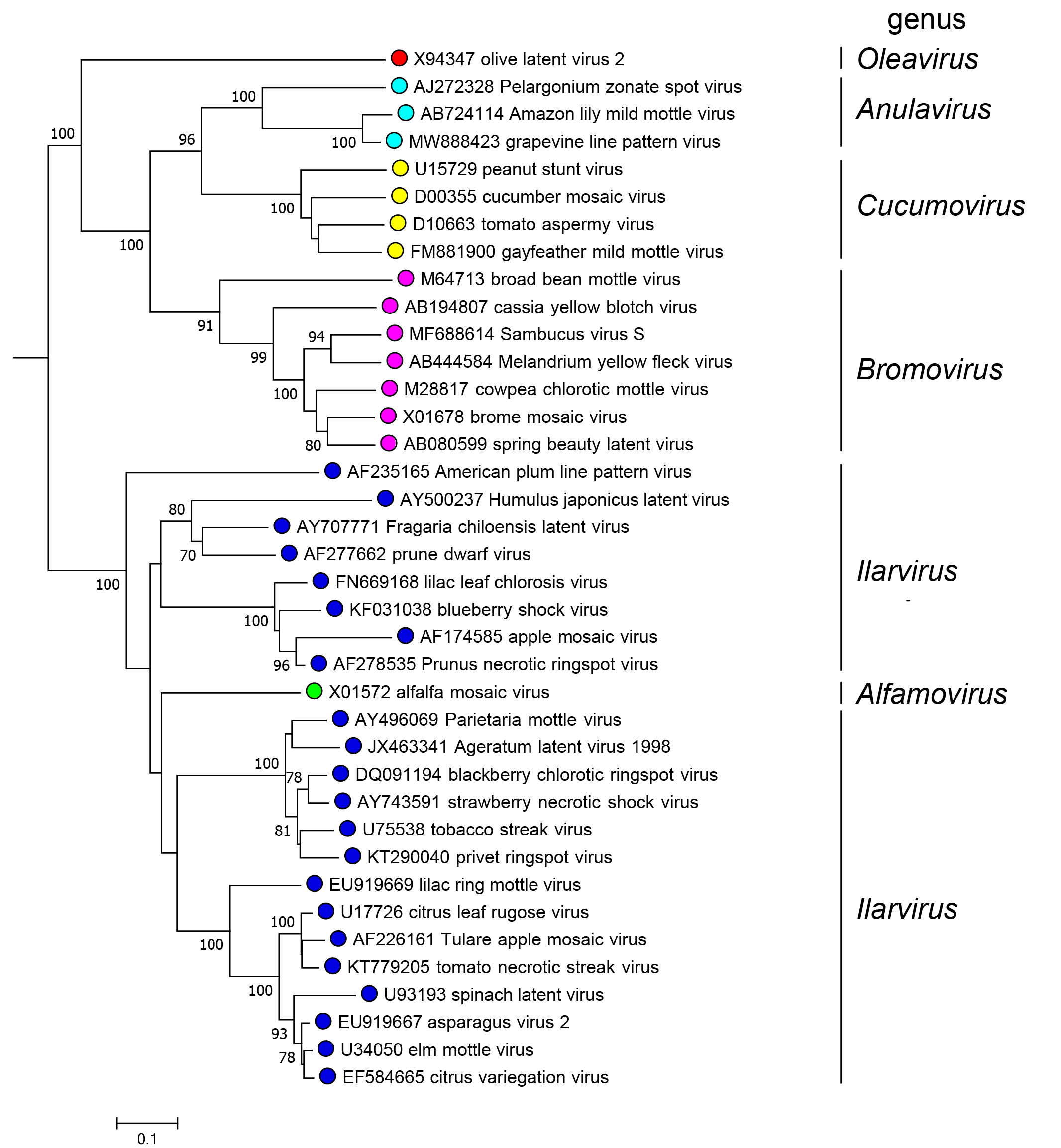 |
| Figure 4.Bromoviridae. Neighbor-joining phylogenetic tree of members of all 38 species in the family Bromoviridae generated from an amino acid alignment of regions spanning the RNA-directed RNA polymerase motifs I to VII (Koonin 1991). Bar = genetic distance. Numbers at nodes = bootstrap values where this was > 70%. This phylogenetic tree and the corresponding sequence alignment are available to download from the Resources page. |
Relationships with other taxa
Bromovirids are members of the “alpha-like” supergroup of viruses, sharing sequence similarity in the 1a protein domains for methyltransferase and helicase activities, and in the 2a protein polymerase domain with plant viruses belonging to the family Virgaviridae (order Tymovirales), and with animal viruses in the family Togaviridae. The 3a proteins of bromoviruses and the 35 kDa protein of members of the genus Dianthovirus (red clover necrotic mosaic virus, RCNMV, family Tombusviridae) form a distinct “family” of movement-associated proteins. Raspberry bushy dwarf virus (RBDV, genus Idaeovirus, family Mayoviridae) and the newly recognized idaeovirus privet leaf blot associated virus (PrLBaV) are similar to bromovirids in genome organization and in the sequence of certain genes. The CPs of RBDV and PrLBaV have RNA-binding domains like those found in the CP of AMV and some members of the genus Ilarvirus and are involved in genome activation (Navarro et al., 2017). Like the members of the genus Ilarvirus, RBDV is transmitted in association with pollen, whereas no natural routes of transmission are known for PrLBaV.
Related, unclassified viruses
| Virus name | Accession number | Abbreviation |
| grapevine angular mosaic virus | AY590305 | GAMV |
Virus names and virus abbreviations are not official ICTV designations.